Chapter 9: Galactic DNA Transport
Chapter 1
Overview of Our Fossils and Their Significance
Paleo Geology of the Site
Arizona was a shallow ocean during both the Cambrian Explosion of Life ~ 500 Ma and during the Permian Explosion of Life ~ 250 Ma.
Shallow oceans were the birthplace of life on earth, and the molecular biology, fluid dynamics, and requisite structures that make this happen are covered in detail in this book.
Tectonic activity eventually lifted Arizona out of the ocean, and subsequent erosion left the ancient ocean floor at the surface in many parts of Arizona. The fossils discovered in these soils not only provide the empirical evidence of how a "birth of life" event happens under the right circumstances, but also provide a glimpse of the scope and magnitude of new life forms created during a quantum speciation event (sometimes called "hyper evolution", a descriptive but technically incorrect term).
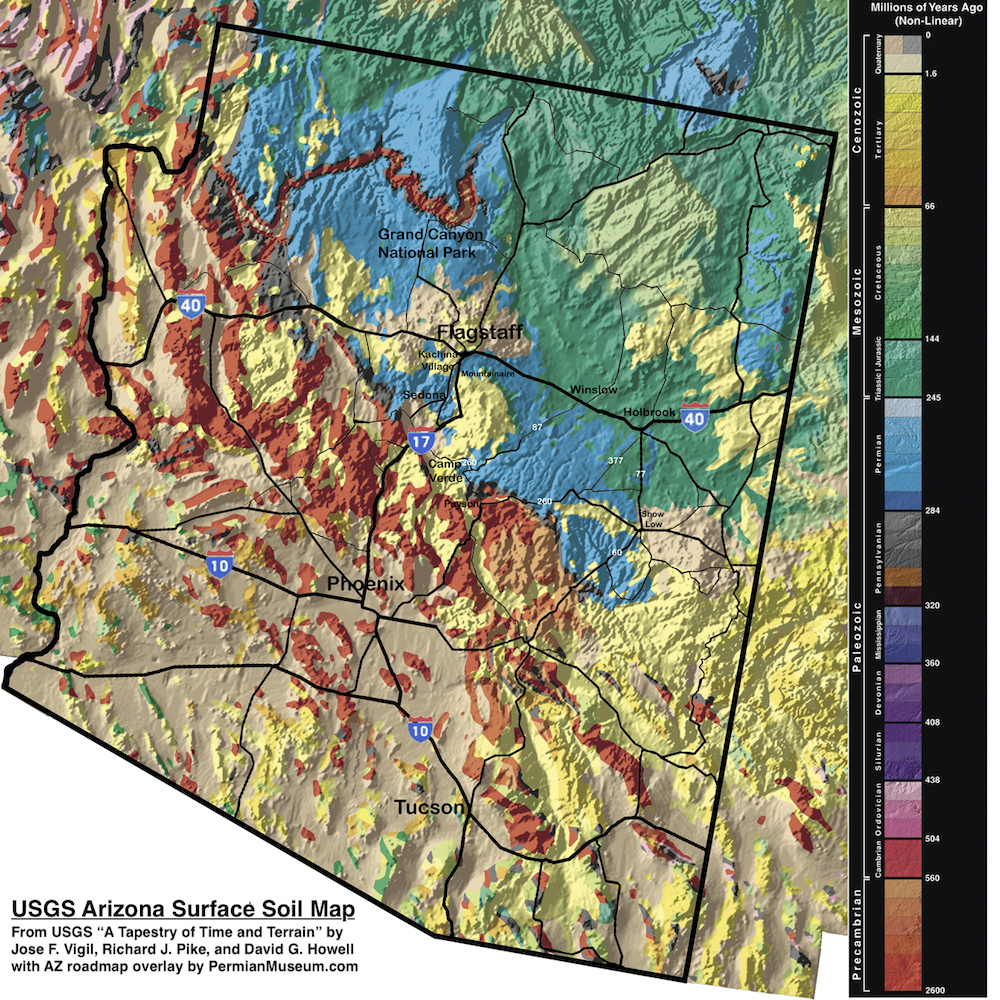
The USGS surface soil map, combined with a current Arizona road map, is shown below. Both Cambrian and Permian fossils can be found at the surface in large parts of Northern Arizona (in red and blue, respectively).
USGS Surface Soil Map of Arizona
The first explosion of multicellular eukaryotic life occurred ~ 500 Ma and is simply known as the Cambrian Explosion of Life. During this time, large parts of Arizona were beachfront property or shallow ocean, as shown in the Map below:
Arizona ~ 500 Ma: The Cambrian Explosion of Life

The Permian Explosion of Life came in 4 pulses. Each started with an extinction event, which was in turn followed by a "Quantum Speciation" pulse that introduced new life forms. The Permian Explosion of Life was much more spectacular than the Cambrian Explosion of Life. The 4 churns of new life forms created eventually introduced a new cast of characters ranging from dinosaurs to birds. While birds live on to this day, the dinosaurs had a great run for almost 200 million years until the next major extinction event came along.
The below graphic shows these pulses, based on data published by Sahney and Benton (Sahney, S and Benton M. J., “Recovery from the most profound mass extinction of all time”, Proceedings of the Royal Society B, 275, 759 - 765 , 2008).
Permian Extinction / Recovery Pulses
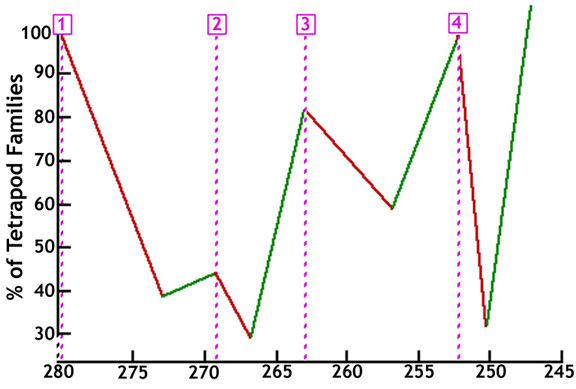
Millions of Years Ago (Ma)
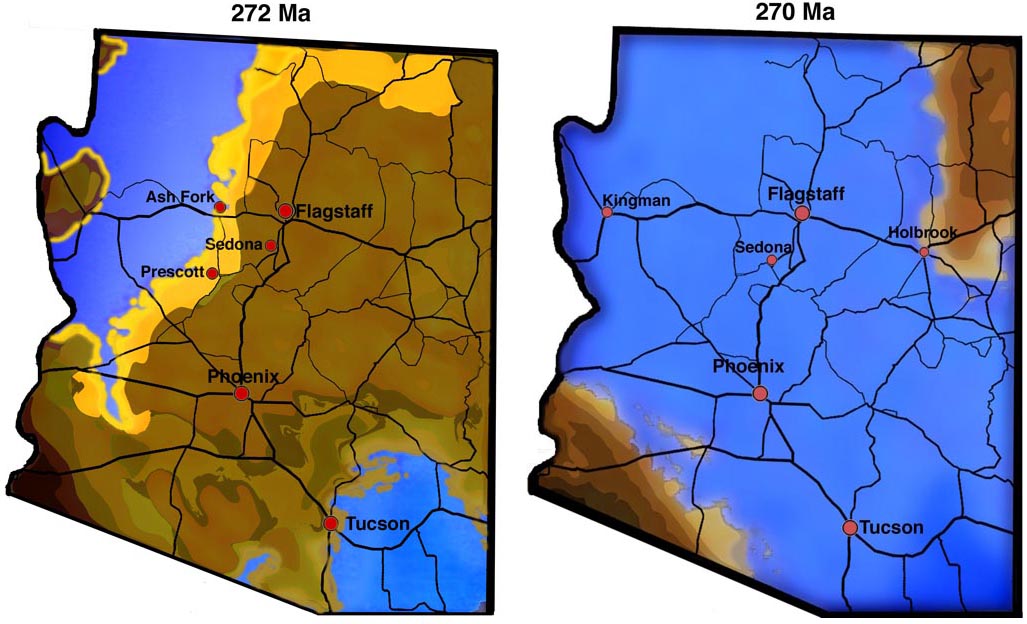
Once again, Arizona was at the right place at the right time to host an explosion of life event. Based on data published by Ron Blakey and Wayne Ranney in "Ancient Landscapes of the Colorado Plateau", Arizona was once again beachfront property or shallow ocean at the start of the Permian quantum speciation pulses.
Arizona ~ 270 Ma: The Permian Explosion of Life
The 35 whole body fossils covered in "On the Origin of Life and Biodiversity", first edition (published in 2014), has been greatly expanded. This online book version allows for continual updates, avoids the prohibitive printing cost because of the large photo and graphic content, and effectively provides a usable textbook free of charge for students and brainiacs on a budget.
Chronological Significance of the Fossils
The Permian Explosion of life 250 Ma was the culmination of the 3 Explosions of Life of earth. It finalized what inhabits the earth today, including land animals.
1)The Prokaryotic (Bacterial) Explosion of Life (3.8 Ba) brought mitochondrial DNA to earth. Mitochondria is a cell’s power plant that stores energy from aerobic respiration (metabolism of glucose). Human mitochondria is bacterial - circular DNA, separate from the nucleus, and its nucleotide sequence points back to a DNA reassortment of rickettsia, rhizobacteria, and agrobacteria.
Physiological life can be defined as a collection of concentration gradients. Their presence means life, their absence means death. It takes energy to maintain concentration gradients. Without the mitochondrial DNA that arrived on earth some 3.8 Ba, life as we know it today would not exist.
2) The Unicellular Eukaryotic Explosion of Life (1.5 Ba): Eukaryotic cells have multiple strands of DNA (not circular) and a single self-assembling lipid bilayer cell wall membrane, both of which facilitate DNA reassortment of eukaryotic cells into new life forms, under the right conditions.
The most notable lineage was the Calcium Secreting Filter Feeders (CSFFs) that lived in colonies and built calcium structures that were able to host DNA reassortment in shallow coastal oceans. CSFFs also brought early bone DNA.
3) The Multicellular Eukaryotic Explosion of Life - 500 Ma (Cambrian) and 250 Ma (Permian):
The earliest known multicellular life forms were microscopic nematodes (roundworms and eelworms) that appeared more than 500 Ma. They are basically a mobile intestine, with a mouth, anus, and gonads (cells that produce haploid cells). They had a protective protein skin and two neurons running the length of the body connecting to muscle that allowed fish like motion. The mobile intestine with gonads went on to become the core blueprint for all multicellular life forms that followed, including humans.
Nematodes provided digestive, reproductive, muscle, nerve, and skin DNA.
The Cambrian explosion of life had unicellular and nematode DNA to work with to generate new life forms by DNA reassortment, followed by 250 million years of evolution.
The Permian explosion had the same input, plus the evolved Cambrian life forms that survived the Permian extinction events. The survivors included sharks, sting rays, reptiles and lung fish, which have also been found and are presented in the book. This allowed a more robust explosion of life that gave rise to the age of the dinosaurs and winged flight.
Life Becomes Aware:
The appearance of a brain cavity that was larger than necessary to house just neurons (shown in the sectioned specimen photo of a winged dragon later in this chapter) allowed for integration and hyper-proliferation of astrocyte cells that interconnect neurons in a way that allows for propagation of the calcium ion wave which hosts the measurable electromagnetic phenomenon underlying consciousness and memory formation.
Neurons make up 97% of a Leech brain. In contrast, neurons only make up 10% of a human brain, and glial / astrocyte cells make up the rest.
The addition of a brain to a mobile intestine with gonads would foreseeably result in a conflicted road toward the development of intellect.
Soft Tissue Fossilization and Some Fossil Photos
Our fossils include preservation of soft tissue as limestone or crystalline forms of limestone. In typical life forms calcium exists as a free ion (Ca++) in solution, or ionically bonded to phosphate in bone, and not as limestone (calcium carbonate or CaCO3).
In many cases, it appears our life forms were instantly turned to stone. To say this type of fossilization is poorly understood is an understatement, however it is not without precedent. It is in respects similar to a case revealed in recently declassified documents where "a source of energy that is still unknown" was able to instantly transform the structure of a living organism "into a substance whose molecular composition is no different from that of limestone." ( source: www.cia.gov/library/readingroom/docs/DOC_0005517761.pdf ). We will refer to this process as "death by limestone fossilization".
A much slower form of soft tissue fossilization as limestone was described in 2005. ( Jozef Kazmierczak and Barbara Kremer, "Early post-mortem calcified Devonian acritrarchs as a source of calcispheric structures", Facies (2005) 51: 554-565.)
The article studied single celled organisms and the authors describe a post-mortem calcium carbonate permineralization that occurs around the organic cell walls. Simplistically, post-mortem bacterial degradation of the calcium rich mucilaginous envelope results in precipitation of fine grains of calcium carbonate, which under pressure of subsequent burial are transformed into crystalline calcium carbonate. The resulting shapes closely match the shape of their organic forerunners.
Conversion of amorphous calcium carbonate into its crystalline forms (Calcite, Aragonite, or Vaterite) depends on several variables including temperature, pressure, time, and the ambient chemical environment ( e.g. ambient Ph, the presence or absence of Mg, the activity of certain microorganisms).
Regardless of how our life forms were fossilized, the soft tissue preservation allows one to see a snapshot of the molecular biology of the time and to infer the DNA that was present at the time, as DNA expression is what generates the biological structures.
The simplest limestone fossilization can be found from temperature alone. The below shows a coral colony cooked, cauterized, and immortalized by the temperature of underlying volcanic activity:
Cooked Coral Colony Fossil

In the side view, the red, avocado shaped bowl at the very bottom is limestone that has been stained by iron that infiltrated from the lava below. Rotating the coral colony rock counterclockwise (front view) reveals a thermal vent. A zoom in on the vent reveals that it is composed of the crystalline form of calcium carbonate, but the rest of the rock is not. Just to the right of the vent, the calcium carbonate is much denser than in other parts of the rock, and it somewhat resembles cement. As will be covered later, external features of some of our fossils are sometimes preserved in the density of the calcium carbonate matrix.
Non mobile life forms, such as coral, would be particularly susceptible to thermal or tectonic death, as they can not run away. Eggs are another example. Eggs that are whole and undamaged would strongly suggest death by thermal activity, or possibly by a high energy event as previously described.
An example of some fairly intact eggs is shown below:
Fossilized Eggs

Sectioned fossilized eggs provide an excellent view of developing life forms within. There are several attributes we use to identify fossilized eggs: 1) the fossil is the shape of an egg (oval or circular), 2) an egg shell is often visible at the periphery in the sectioned specimen, 3) if there is a developing life form inside, there is often a surrounding calcium powder filled space around the life form as the egg yolk has been consumed and 4) the egg shaped fossil is often attached to a substrate, and found with other eggs and life forms in proximity.
A few photos are presented below to provide a better idea of what the fossilized eggs look like.
The example below shows the external view of an egg that has been eroded to reveal the egg shell and contents inside.
Erosion Exposed Egg Shell and Egg Contents

The next example is a sectioned egg that shows an indeterminate life form inside that has eaten part of the yolk, which leaves a space surrounding the life form.
Void Surrounding the Life Form as Yolk is Consumed

Near some of the egg nest areas there are also fossilized remains of intact bird like life forms, indicating the non destructive form of death happened over a large area. The bird fossils tend to indicate the more instantaneous death and fossilization process. A picture of some of the larger bird forerunners is shown below:
Fossilized Birds

A sectioned specimen of a small bird like life form shows the same crystallization pattern, with the life form entombed in a calcium carbonate matrix. Interestingly, the life form appears to be glancing back a something that caught its attention, just before being turned to stone. This is consistent with the visible component of the high energy release associated with the death by limestone fossilization event, as described by the survivors that were protected by trees.
Sectioned Small Coastal Bird

However, it is also possible they may have been killed by a thermal event. A bird may not flee a thermal event, such as lava heated water, because of the neurology of the time. Sectioned specimens reveal that most of the newly created life forms did not have a brain or even neurons for sensing temperature. The basic arrangement was an optic, olfactory, or auditory neuron connected directly to a motility group.
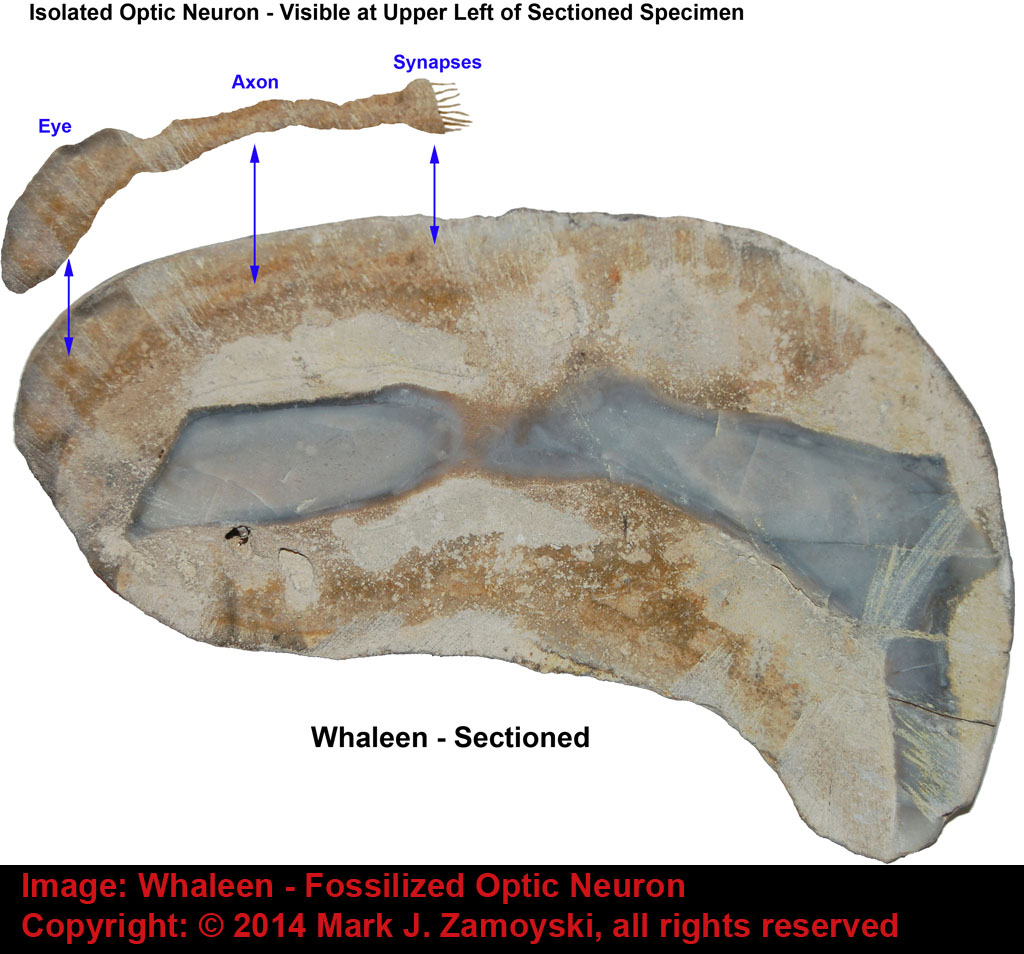
As an example, Whaleen had a single optic neuron that connected directly to a motility group (its tail).
Illustration of Specimen Called Whaleen:

Photo of External View of Whaleen with Matrix Removed:

Sectioned View of Whaleen Reveals Single Fossilized Optic Neuron:
Dino-Seal is the most neuronally advanced brainless specimen, with 3 neurons ( Olfactory, Optic, and a possible vibration sensing neuron), each connecting to a motility or directional guidance group, but no brain.
Sectioned View of Dino-Seal Reveals Three Fossilized Neurons:

Accordingly, given the neurology of the time (i.e. absence of pain sensing neurons or a brain), the life forms would have been cooked by the thermal event, without even realizing what was happening, and subsequently fossilized by the thermal event. It is also possible that life forms that had the ability to sense and flee thermal danger existed, and are not in the collection because they were able to avoid such dangers.
We did not start finding evidence of life forms with a brain until we began processing some of the larger specimens.
The Winged Dragon fossil is an example.
Winged Dragon with Oversized Brain Cavity

Neurons make up 97% of a Leech brain but only 10% of a human brain. Glial / astrocyte cells make up the rest in a human.
The appearance of a brain cavity that was larger than necessary to house just neurons allowed for integration and hyper-proliferation of astrocyte cells that interconnect neurons in a way that allows for propagation of the calcium ion wave which hosts the measurable electromagnetic phenomenon underlying consciousness and memory formation.
As for the tissue preservation aspect of the discussion, it should be noted that the sectioned specimen of Whaleen above shows what appears to be gestating progeny in an isolated sack in the central food lumen (more typical of a nematode). The progeny have been preserved in crystalline form, versus the rest of Whaleen, which has been preserved as hardened calcium carbonate. This aspect of the soft tissue preservation is not yet understood, as undigested stomach contents in specimens such as Zamoyski dragon are not preserved in crystalline form, but as hardened calcium carbonate, just like the remainder of the life form:

The undigested stomach contents (Goby Shark) found in the central food lumen of the Zamoyski dragon are not preserved in crystalline form, but as hardened calcium carbonate, like the rest of the Zamoyski Dragon. In contrast, the gestating progeny inside Whaleen are preserved in crystalline form, while the remainder of Whaleen is preserved as hardened calcium carbonate.
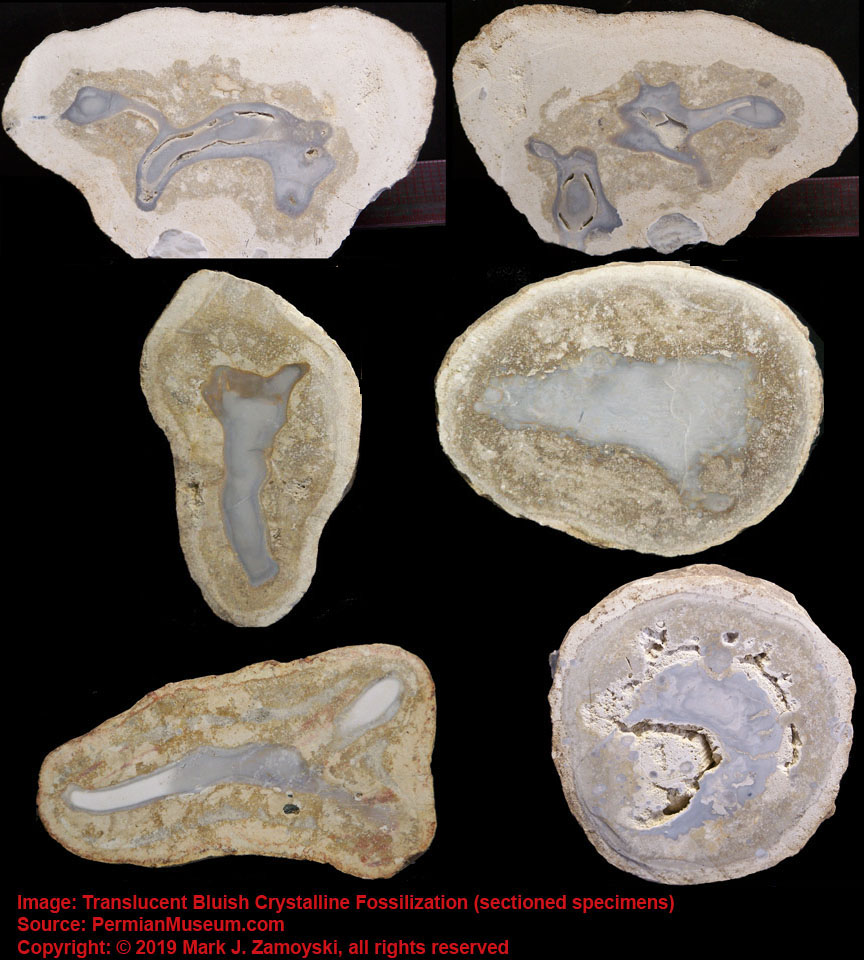
Further complicating an understanding of the fossilization process is that roughly 15 - 20% of the life forms are whole body fossils that are preserved as a translucent bluish crystal. They are similar to the progeny inside Whaleen, except that the entire life form is preserved as a translucent bluish crystal. Examples:
Examples of Entire Life Form Fossilized as a Translucent Bluish Crystal
And if that was not enough, in early 2020 we discovered a third, very rare, and much more spectacular form of fossilization we call "the resin embeds". We do not understand the geology that created them, but they appear to be life forms embedded in a yellowish crystal matrix. An example of one such specimen is shown below:
Example of "Resin Embed"

Observations about the appearance of our fossils:
1) The full crystallization of larger life forms is extremely rare and external features are rarely preserved well, while internal features are more commonly preserved well.
2) Sometimes external features are preserved in the density of the surrounding matrix, and as such progressively harsher removal methods can destroy these features.
3) When external preservation does occur, it is often much better on one side and not the other. This is possibly due to which side was on the bottom during the postmortem fossilization process. It may alternatively be related to manner of death, as some more specimens appear to have compression on one side and distention on the opposite side (e.g. eye crushed in on one side and the eye on the other side bulging out of its socket). This would be consistent with the life form being killed by a high pressure event from one side.
4) The internal preservation visible by sectioning a specimen (i.e. cutting it in half) is so good some times that it reveals anatomical features such as neurons, undigested stomach contents, and even gestating progeny. The internal preservation can typically reveal the general external morphological shape of the life form much better than removing the external matrix.
5) The key to seeing many of the internal structures in photos depends heavily on the light source used. Light sources that penetrate the crystal, with minimal surface reflectivity, provide the best photos.
Calcium Secreting Filter Feeder (CSFF) Photos
The Permian coastal ocean floor was covered with hard skeleton labyrinth structures built by single celled organisms called Calcium Secreting Filter Feeders (CSFFs).
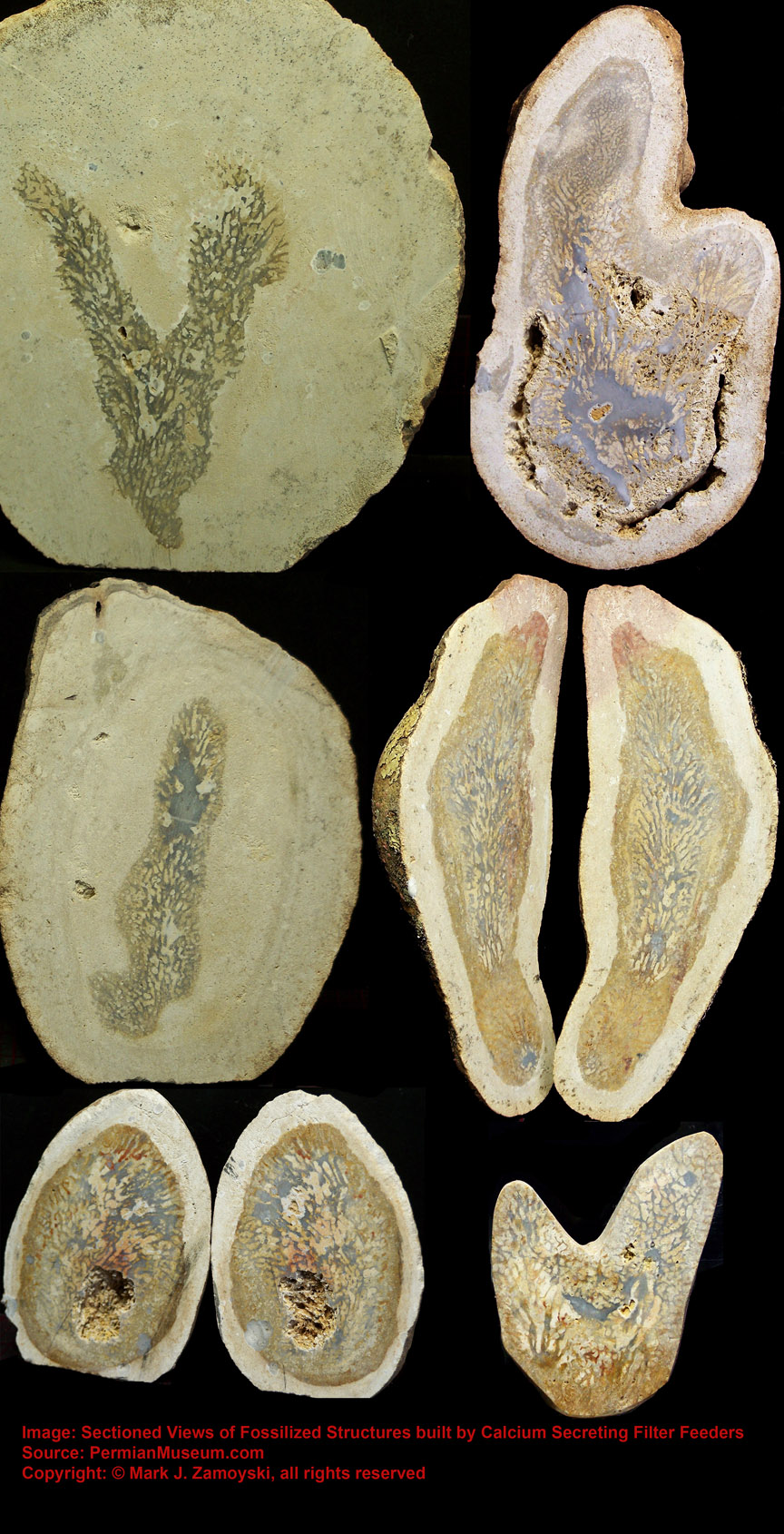
They are the most prevalent, best preserved, and most significant fossils in our collection. Most have been turned into a crystalline form of calcium carbonate and are encased in a calcium carbonate matrix. A photo of the variety of these CSFFs is shown below. The top and bottom rows are sectioned specimens. The middle row shows the external view of exposed specimens.
Calcium Secreting Filter Feeders (CSFFs)

These are the most significant fossils in the collection because they are capable of hosting "Quantum Speciation", or the abrupt and abundant appearance of new life forms without antecedent lineage, through a process that can best be described as "Grab Bag Genetic Reassortment".
An example of a cracked open rock revealing the external features of such a structure shown below:
Permian Protopharetra - Exposed Exterior

Sectioned specimens are even more spectacular and provide an entirely new narrative on the origin of life and biodiversity on earth.
Permian Protopharetra - Sectioned Specimen
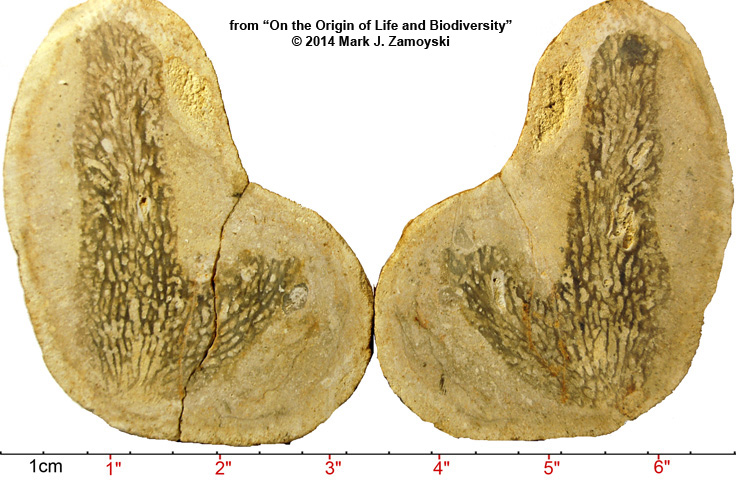
Author is extremely grateful to Dr. James T. Sprinkle of the University of Texas at Austin, who took the time to look at the picture and identify a potential Cambrian counterpart. A smaller version of the specimen above first appears in the fossil record around 500 ma at the start the Cambrian Explosion of Life (Boardman, Fossil Invertebrates, 1987, p.114 Figs. A and B ) and was called a Protopharetra. It appears here again during the Permian Explosion of Life, and author has named this larger version the Permian Protopharetra.
Protopharetra structures were built by Archaeocyathans, the first unicellular eukaryotes that lived in colonies and secreted calcium carbonate as a skeletal matrix. These organisms were filter feeders, channeling moving ocean water through the hard skeleton labyrinth, to obtains suspended nutrients.
On earth, new viruses are created by genetic reassortment of existing viruses in co-infected cells.
Similarly, the evidence presented on this website will show that new life forms on earth can be created by DNA reassortment of existing life forms, hosted in CSFF structures.
A cubic inch of coastal water contains more than 15 million suspended cells, from a large variety of life forms. CSFFs channel this water, shearing open cell membranes to release the DNA, proteins, and other intracellular contents to the feeding colony below.
In 2014, we published sectioned CSFF specimens that revealed an unintended consequence of these structures was their ability to host a ≥grab bag≤ DNA reassortment process. This would result in creation of new life forms, such as those that spawned the age of the dinosaurs.
Combining fluid dynamics with molecular biology reveals how these structures are capable of hosting this "grab bag" DNA reassortment process. A detailed presentation of several of the CSFF structures, and a review of how they fulfill the requirements necessary for genetic reassortment, under known principles of fluid dynamics and molecular biology, are included on the website:
Based on the DNA pool available as input into the process, there would be several expected outputs of such a process. They would include new life forms that appear to be quantum leaps backward in evolution, others that appear to be quantum leaps forward in evolution, others that have duplications or redundant features, others that are clear forerunners of the dinosaurs, as well as bizarre, never before seen new life forms. We now have fossil evidence for all of these predicted life forms.
The DNA pool available for input into the process is summarized in "The Input" section of Chapter 4:
The list of expected new life forms from such a process, given the available input DNA, is listed at the beginning of the "Output Specimens" section of Chapter 4 and the actual fossil evidence of all of the new life forms is presented in the balance of Chapter 4. A photo gallery of the Indeterminate / Unknown Specimens is in Chapter 6.
The other huge empirical evidence that this process actually occurred are the re-speciation timelines observed after an extinction event. They are too short for evolution, but more than adequate for quantum speciation by grab bag DNA reassortment. The below graphic depicts these pulses, based on data published by Sahney and Benton (Sahney, S and Benton M. J., “Recovery from the most profound mass extinction of all time”, Proceedings of the Royal Society B, 275, 759 - 765 , 2008)
Permian Extinction / Recovery Pulses
Millions of Years Ago (Ma)
The Extinction / Recovery Pulses are also consistent with two other expectations.
First, the 4 extinction pulses that occurred during the Permian period were not complete extinction events, as a small number of life forms survived the extinction. This means that "survivors" would also be present at the same time "newbies" were being created. The "survivors" were likely spawned 500 million years ago (newbies at that time) by the grab bag DNA reassortment process and then had 250 million years of evolution under their belt before the Permian period extinction pulses came along. As such, survivor DNA could be expected to be more complex, and would also be available for the grab bag DNA reassortment process. This could be expected to make the Permian Explosion of Life much more robust than the Cambrian Explosion of Life, as the deck was stacked with the superior "survivor" DNA during the Permian Explosion of Life.
Second, a well populated, established ecosystem could be expected to be highly detrimental to survival of new life forms. As interesting as a new life form may be, in a well populated ecosystem it would merely be a snack for one of the numerous established predators. Basically, the steeper the extinction pulse, the steeper the recovery pulse. While the chart shows terrestrial tetrapod families, the end permian extinction event wiped out 96% of marine life. This created a fairly clean slate for the survival of new life forms spawned during the re-speciation pulses, some of which would also eventually migrate onto land.
The churning out of new life forms over 4 quantum speciation pulses resulted in the introduction a completely new cast of characters which ushered in the age of the dinosaurs and winged flight. These were life forms with no antecedent lineage. This is what DNA reassortment does. This is something evolution / selective advantage can never do, as evolution requires an antecedent lineage as a starting point, and selective advantage favors small improvements over long periods of time.
While the appearance on new life forms is sometimes referred to as "Hyper-Evolution", that is a descriptive, but technically incorrect term. The underlying process is "Genetic Reassortment" that starts with a single newly created cell in a nutrient rich environment capable of hosting its growth and division (similar to an egg). The underlying molecular biology of this process, and the fluid dynamics of why shallow oceans are ideally suited to host this process, are covered in the next chapters.
Chapter 2
A Brief History of Life on Earth

The universe is estimated to have originated from the big bang some 13.7 billion years ago (Ba).
The earth formed around 4.6 Ba, with the peak of the meteoroid impacts occurring around 3.9 Ba.
The Oceans formed around 3.8 Ba.
Atmospheric oxygen began appearing around 2.5 Ba, and was close to current levels by 1.5 Ba.
Three explosions of life occurred in earth’s history.
The Prokaryotic Explosion of Life
Unicellular Prokaryotic Cells (aka bacteria): Prokaryotes first appeared 3.5 - 3.8 Ba. Chemical traces of prokaryotic cells date back to 3.8 Ba, or 0.1 Ba after the end of the asteroid impacts on earth. Fossil evidence dates back to 3.5 Ba.

Cyanobacteria were among the first prokaryotic cells to appear and can live in anaerobic or aerobic environments. They obtain their energy by photosynthesis, taking an electron from water and releasing oxygen as a waste product. They are believed to have created our oxygen atmosphere.
Cyanobacteria also convert carbon dioxide into carbohydrates, providing a food source, and make the enzyme nitrogenase, which fixes nitrogen into a form that can be absorbed by plants and used in the synthesis of proteins and nucleic acids.
Mitochondria is a cell’s power plant that stores energy from aerobic respiration (metabolism of glucose) and mitochondrial DNA is bacteria's most direct contribution to life on earth.
Human mitochondria is of prokaryotic origin: its DNA is separate from that of the nucleus, is circular (bacterial), and its nucleotide sequence analysis points back to a DNA reassortment of rickettsia, rhizobacteria, and agrobacteria ( Alberts et. al., Molecular biology of the Cell, Third Edition 1994).
Physiological life can be defined as a collection of concentration gradients. The presence of concentration gradients means life. The absence of concentration gradients means death. It takes energy to maintain concentration gradients. Without the prokaryotic mitochondrial DNA that arrived on earth some 3.8 Ba, life as we know it today would not exist.
Prokaryotic cells have a tough triple layer cell wall, and can thrive near volcanic vents 3,500 feet below the ocean surface and live two miles deep in soil at pressures of 5,000 PSI. Bacteria have circular DNA. They may have a flagella or whip like tail for propulsion, and can aggregate in colonies that function as a unit.
The prokaryotic explosion of life was so successful that today bacterial biomass on earth exceeds that of all plants and animals combined. One gram of soil contains 100 million to 1 billion bacterial cells. Coastal oceans contain 1,000,000 cells per ml.
The Unicellular Eukaryotic Explosion of Life
Unicellular Eukaryotic: Eukaryotes first appeared 1.5 Ba. Evidence indicates they hail, in whole or in part, from prokaryotic cells.
They have a soft single layer cell wall membrane, which is basically one of the three layers of a bacterial cell wall. They have multiple strands of linear DNA that is contained in a membrane bound compartment called the nucleus. Eukaryotic mitochondria is of prokaryotic origin: its DNA is separate from that of the nucleus, is circular (bacterial), and its nucleotide sequence analysis points back to early bacterial origins.
When your favorite detective show talks about matching a suspect's mitochondrial DNA to mitochondrial DNA found at the crime scene, what they are really saying is “Which combination of bacteria does our suspect hail from, and does that combination match the one found at the crime scene?”.
The fact the mitochondrial DNA is from several different prokaryotic cell types is also proof that DNA reassortment does happen.
The eukaryotic multiple strands of DNA that code for the rest of the life form and the single self-assembling lipid bilayer cell wall membrane readily facilitates DNA re-assortment into new life forms under the right conditions.
The most notable eukaryotic cells were a lineage of Calcium Secreting Filter Feeders (CSFFs) that were able to host DNA reassortment when combined with the fluid dynamics of shallow coastal oceans. They also provided the DNA for bones, which in turn allowed for Calcium/Phosphorous/Growth Factor mediated multicellular life forms.
The Multicellular Eukaryotic Explosion of Life
A multicellular life form is a collection of eukaryotic cell types that live and function as a unit. In a multicellular life forms, each cell contains the complete DNA to code for all of the cell types, but expresses only its subset of that DNA that makes it a specialized cell type. This simple fact indicates a completely new multicellular life form can only arise at a unicellular level. Multicellular life as we know it starts from a single cell with the complete DNA code, and that cell then goes on to grow and divide into the trillions of cells and hundreds of specialized cell types coded for by the DNA contained in that first single cell.
Nematodes (roundworms and eelworms) are the oldest known multicellular eukaryotic life forms, with fossils dating back to the Cambrian period (543-490 Ma) and because many are microscopic and transparent the estimates of their first appearance date back to Pre - Cambrian times.
CDC Photo of Intestinal Nematodes
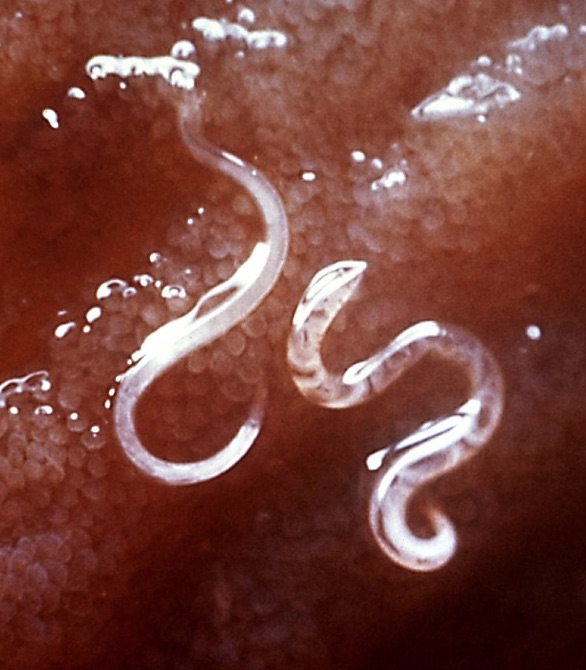
Nematodes have a tubular digestive system, with a mouth at the front and anus at the back. They have internal male, female, or hermaphroditic reproductive organs, where eggs are produced and pass out the digestive tract into the environment. They are invertebrates that have an exoskeleton made up of a hard, but flexible, cuticle on the outside (secreted by their epidermis). Nematodes have 2 nerves, one on the back and one on the belly, that run the length of the nematode. The nerves connect to muscles attached to the cuticle, allowing for side to side (fish like) motion.
Nematodes are basically a mobile intestine, with a mouth, anus, and gonads (glands capable of producing haploid cells) and effectively served as the core blueprint for all multicellular life forms that followed, all the way through to humans.
Nematodes also brought digestive, reproductive, skin, nerve, and muscle DNA to the subsequent DNA reassortment parties. The same neuron related proteins in transparent roundworms such as Caenorhabditis elegans are still found in humans today.
The Nematode explosion of life was almost as spectacular as the prokaryotic explosion of life, with around 1 million nematodes per square meter of soil today. Nematodes are also the most prevalent life form on the ocean floor.
The appearance of Calcium Secreting Filter Feeders during both the Cambrian and Permian Explosions of life, which were capable of both hosting Grab Bag DNA Reassortment, as well as likely providing their DNA for the first "Boned" life forms, created an unimaginable spectrum of new life forms (the process is covered in Chapter 3 and the empirical data (fossil record) corroborating the process is presented in Chapter 4). Only a small number eventually made it into earth's playbook of life.
The Cambrian explosion of life had prokaryotic, eukaryotic, and nematode DNA to work with to generate early new life forms, followed by 250 million years of evolution to improve them.
The Permian explosion had the same 3 primitive DNA sources plus more advanced Permian extinction survivors such as sharks, lung fish, sting rays, and reptiles as input for the second round of DNA reassortment. This allowed for a more robust explosion that spawned the age of the dinosaurs and winged flight.
A few examples of the life forms created during the Permian Explosion of Life:

The more robust Permian Explosion of life introduced the age of dinosaurs and birds, which dominated the earth from around 250 Ma to 66 Ma, when the global cooling from an asteroid impact (K-T extinction event) wiped out terrestrial life forms weighing more than 25 Kg (55 lbs.).
The dramatic climate change caused by the K-T asteroid was relatively short. The freezing temperatures reversed after 3 years (Brugger, Julia; Feulner, Georg; Petri Stefan (2016) "Baby, its cold outside: Climate model simulations of the effects of he asteroid impact at the end of the Cretaceous" ) Geopysical Research Letters. 44(1): 419-427) and returned to normal within decades (Whittle et. al., 2019, "Mass Extinction", Dryad Digital Repository) .
The Paleocene period ( 66 - 56 Ma) that followed saw a tropical, greenhouse climate with forests worldwide and no polar ice. Global average temperatures were ~ 24 C (76F) or 10 C warmer than the 1951-1980 average of 14C (57F), and CO2 levels were around 500 ppm versus 400 ppm today. The Eocene period ( 56 -34 Ma) that followed got even better, after 2,500 -4500 gigatons of carbon were released into the atmosphere, with the highest measured levels of CO2 reaching 4,000 ppm, and average temperatures rising by 5 - 8 degrees C (9 -14 F), which put the temperature around 15C ( 28 F) warmer than today. By the end of the Eocene, CO2 levels had decreased to around 800 ppm.
This climate is what hosted the rise of our forerunners. Primates are believed to have arisen from small terrestrial mammals and the first small monkey like life form (Haplorhini) appeared 63 Ma, or 3 million years after the K-T extinction event. Other small monkeys and apes appeared around 40 Ma. The great apes (gorilla, orangutan, chimpanzee, bonobo) appeared around 17 Ma. The Homo ("man" in Latin) genus first appeared ~ 2 Ma with Homo Erectus, which went on to spawn the Homo Neanderthalensis and Homo Sapiens (anatomically modern humans), which first appeared ~ 300,000 years ago. The earliest Homo Sapien fossils found so far date back to ~ 315,000 years ago.
That brings us to the latest multicellular explosion of life.
The Human Explosion of Life - A First Order Logarithmic Progression Problem
It took humans ~ 315,000 years to get through the first 30 population doublings to get to 1 billion people in 1800. The next population doubling took 130 years, with 2 billion people in 1930. After that, things really went off the rails. Scientific advances in medicine and agriculture all resulted in a further reduction in the population doubling time. The next population doubling only took 44 years with 4 billion people in 1974. The latest population doubling has taken 46 years, with ~ 8 billion people by the end of 2020.
The carrying capacity of an environment is defined as the maximum population size of a biological species that can be sustained in that specific environment, given the food, habitat, water, and other resources available.
In 2001 the UN stated the estimates of the carrying capacity of the planet for humans varied widely, with a median estimate of 10 billion, and the top of the range at 16 billion.
It is important to note that the carrying capacity estimates were based on current baseline temperatures. Warm temperatures increase food production and carrying capacity. Cold temperatures reduce carrying capacity, and based on what we know about earth's orbital cycles (as corroborated empirically by the Vostok ice core data), global cooling will once again return:
The Return of Orbit Related Cooling: Reduced Carrying Capacity

2025 UPDATE: Because of COVID / mRNA VAX reduced fertility rates, the UN has adjusted its estimates of when we will hit the 10 Billion median estimate of carrying capacity to the year 2061 and the World Bank estimates 2050.
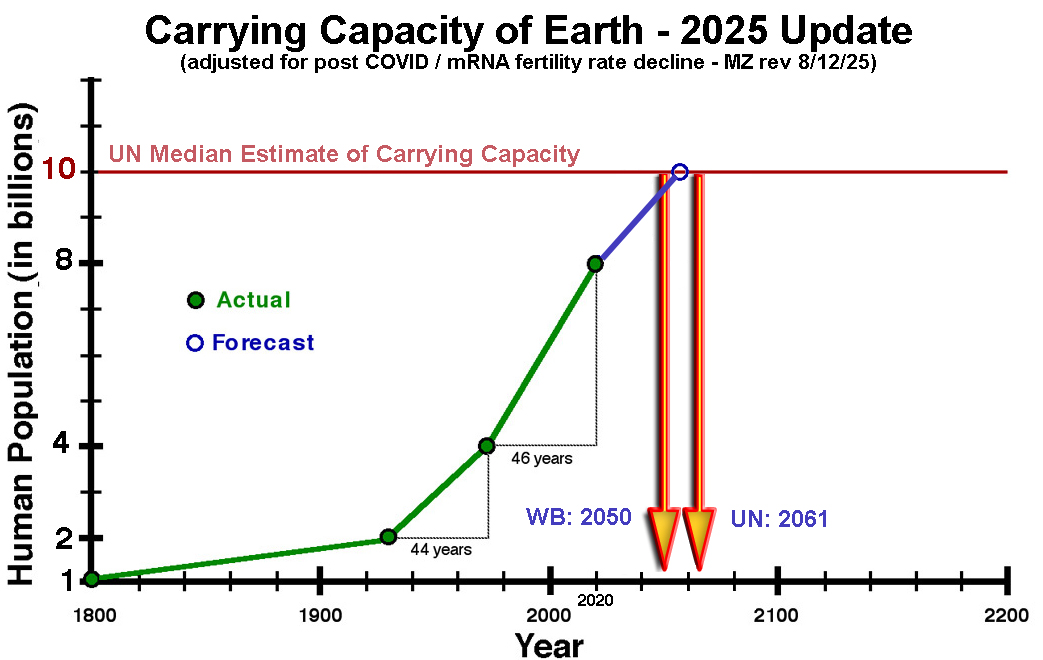
CAVEAT: The full impact of the mRNA VAX death and disability, for 5+ Billion people, is not yet known.
From a molecular biology perspective, the foreseeable death, disability, and fertility, resulted from 3 simple reasons, and it is not over yet.
1) NON-SELF PROTEIN INDUCED CELL DEATH
The COVID virus only infects cells with ACE2 receptors, makes them produce non-self proteins, and those cells are then killed by your immune system (via MHC1).
In contrast: mRNA is delivered by the LNP to all 210 cell types, makes them produce a non-self protein,
and those cells are then killed by your immune system (via MHC1).
This Kills blood vessel (endothelial) cells that results in clot formation, long term heart and blood vessel damage (scarring and plaque), myocarditis, pericarditis, death, and elevated future death risk.
Kills myelin producing cells, resulting in nerve damage from de-myelination.
Kills brain cells, esp. in the cortex and cerebellum, used for balance, coordinate movement, attention, learning, and emotion regulation. Affects can range from "Biden Brain" to "mRNA Reavers" (Throp et al : violent behavior 80 X more likely, homicidal ideation 25 X more likely, etc…)
Kills developing fetus cells, resulting in a ~ 10% miscarriage rate.
Kills reproductive organ cells, resulting in infertility in both males and females. Duration yet unknown.
Immune systems cells themselves can also get infected and be killed, and the thymus is known to be damaged, resulting in mRNA acquired immune deficiency - as of yet unknown duration.
and the list goes on for the other cell types …
2) FULLY ACTIVE "SPIKE PROTEIN" ACTIVITY
A internalized Spike protein binds with P53 and BRCA (disclosed by Dr. Ryan Cole), which suspends DNA damage checkpoints, both accelerating existing cancers and facilitating development of new cancers. It is not uncommon for viruses to do this (e.g. HPV) and the affects are long term.
3) "BADJUVANTS"
The carcinogenic DNA template contaminants will take time for their affects to fully manifest.
The last minute switch to add SV40, a know carcinogen, will also increase cancer rates.
The untoward mRNA coding for amyloid and prions (per Dr. Robert Malone) has yet to be explained. The blood vessel cell death mentioned above evokes a natural fibrin formation clotting process, that with amyloid now added, may account for the string clots observed.
We have not yet seen the full extent of the mRNA induced population decline.
And with recent announcement of a 100% mortality rate "Gain of Function" Avian Flu developed, the carrying capacity problem may be a moot point.
Chapter 3
Quantum Speciation by Grab Bag DNA Reassortment
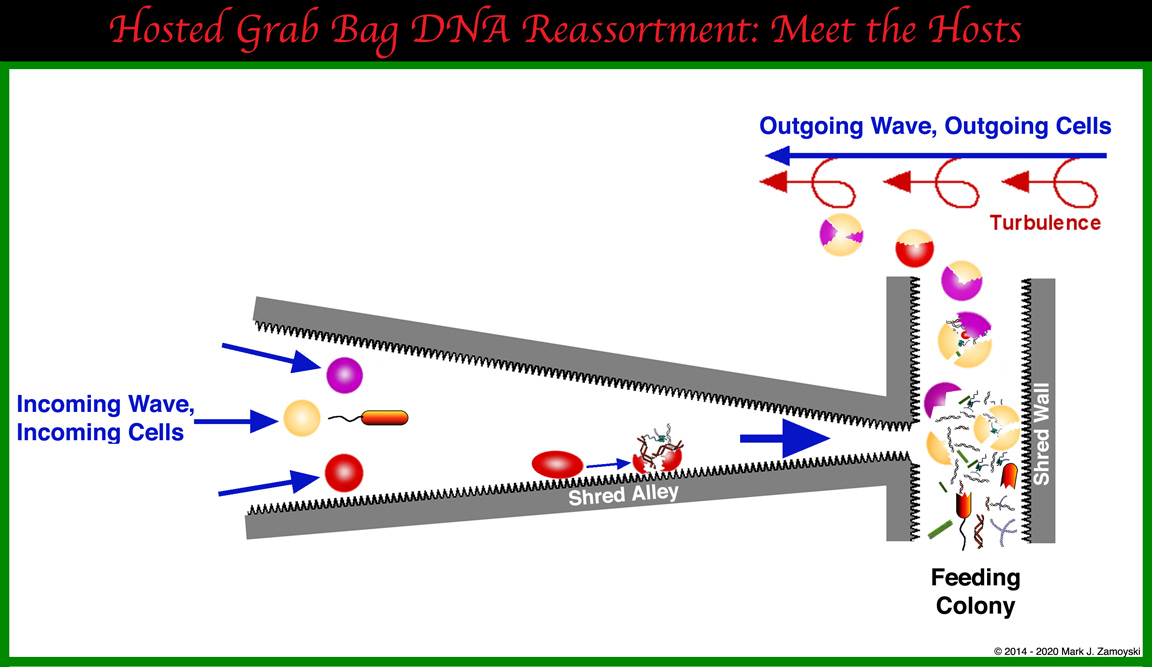
In this section, we will evaluate hard skeleton structures built by a class of single celled organisms collectively called Calcium Secreting Filter Feeders (CSFFs). In particular we will be looking for the ability of these structures to host a DNA reassortment process capable of generating new life forms. A brief synopsis of the relevant related molecular biology and fluid dynamics is presented first, to lay the groundwork for an understanding of the rest of this section.
Relevant Molecular Biology
Cell Walls
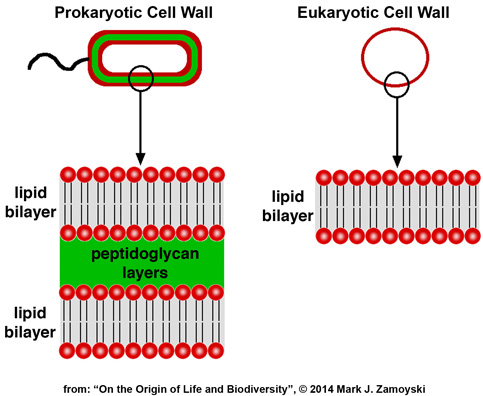
Prokaryotic cells have a rigid, triple layer cell wall structure. Eukaryotic cells have a flexible, single lipid bilayer membrane that is a subset of a prokaryotic cell wall, shown diagrammatically below:
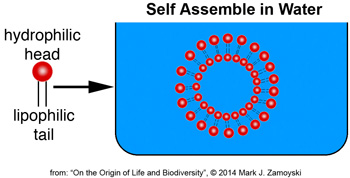
Lipid bilayers are made up of molecules that have a water loving head (hydrophilic) and lipid loving tail (lipophilic). When placed in water, they self assemble to form compartments. Likewise, if a lipid bilayer of the prokaryotic wall shown above was scraped off in water, it would self assemble into a lipid bilayer compartment.
Prokaryotes lack internal membrane bound compartments. Eukaryotes use internal membrane bound compartments (nucleus, mitochondria, Golgi apparatus). The membranes are also made of these lipid bilayers.
DNA and DNA Expression
DNA is the blueprint for proteins. DNA expression means synthesis of proteins from that blueprint. Synthesis of proteins in eukaryotic cells is achieved by a process that involves 1) Transcription of DNA into mRNA in the nucleus, 2) Transport of the mRNA strand to the ribosome (made up mostly of rRNA ), and 3) complimentary base pair binding of tRNA with an attached amino acid, whereby the mRNA strand is translated into a protein. Proteins make up 60% of a cells dry mass and determine what a cell does.
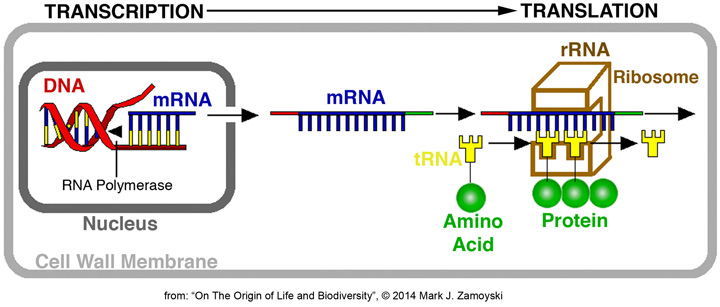
DNA expression is regulated by numerous pathways, including endocrines produced by distant cells (cell signaling).
DNA Efficiency
A simple measure of genomic efficiency can be made by comparing how many proteins are synthesized per million base pairs of DNA.
The cells with the best genomic efficiency could be argued to be the most advanced.
Prokaryotic cells have a single circular DNA chromosome. Some have two.
Eukaryotic cells have linear DNA. Humans have 46 strands joined as 23 chromosomes. The notable exception is mitochondrial DNA, which is circular and resides outside of the nucleus.
The human linear fragments look like a debris field when compared to a simple circular DNA chromosome.
So how does today’s linear eukaryotic DNA compare to the 3.8 billion year old circular prokaryotic DNA?
The genomic efficiency of ancient prokaryotic cells (from “On the Origin of Life and Biodiversity”, © 2014 Mark J. Zamoyski, Appendix A) versus a human eukaryotic cell (DOE, Human Genome Project, Oct. 2004 findings) is summarized below.
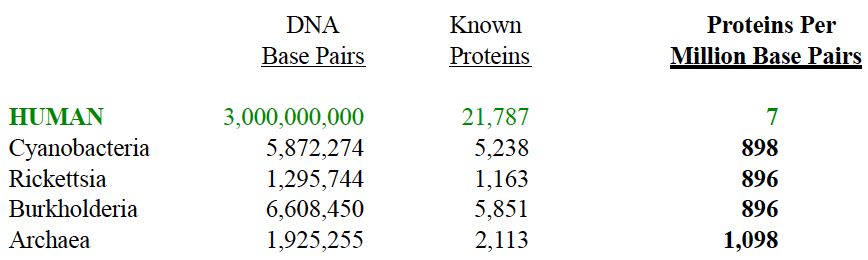
Well isn’t that interesting.
The supposedly superior human eukaryotic cell cranks out only 7 proteins per million DNA base pairs versus bacteria that crank out around 900 proteins per million base pairs. The 3.8 billion year old cyanobacteria’s circular DNA is some 130 times more efficient than the linear human DNA. Archaea, the oldest known prokaryotic cell, is 156 times more efficient.
The human genome project also revealed that 98% of human DNA is non-coding (i.e. not used). We are basically a genetic wasteland, with a few good sequences. The eukaryotic DNA not only looks like a debris field, it is one.
DNA Reassortment
If one desired to create new eukaryotic cells with enormous potential biodiversity, it would require only three conditions.
1) Cells aggregated in close proximity to each other in water:

2) Shear forces or structures capable of rupturing cell membranes:
3) A confined space where the spontaneously reassembling lipid bilayers could effectively encapsulate a batch of the ambient genetic slurry.
The “grab bag” or random DNA reassortment process could be expected to generate cells with much lower genomic efficiency than the cells one started out with, as well as having much unused DNA.
Although both prokaryotic and eukaryotic cells could be used as input into the process, only eukaryotic cells would emerge as output of the process, because of the ability of their cell membranes to self assemble.
The resulting eukaryotic cells could also be expected to have membrane bound compartments inside the main cell wall membrane compartment.
Relevant Fluid Dynamics
Coastal oceans have around 1,000,000 suspended cells per ml of water. To obtain the intracellular nutrients, such as proteins and nucleotides, the cell wall would need to be sheared open. This is the presumed motive for channeling water through the hard skeleton labyrinth structures built by the CSFFs.
But are these structures also capable of creating cells with reassorted DNA?
Understanding a couple of fluid dynamics concepts is necessary to complete the picture.
Water Velocity Amplification: For a given flow (e.g. in cubic mm / sec) coming in from a source (pipe, ocean etc...), water velocity increases exponentially as the water passes through a confined space. The reason is that flow (Q) equals the velocity (V) times the cross sectional area (A) or Q= VA. The area (A) of a circle is A= 3.14 X R2 where R is the radius. For a given Q, a reduction in radius results in an exponential reduction in area (i.e. R2), which in turn requires an exponential increase in velocity to maintain equality.
A simple example of this is a fire hose nozzle attached to a fire hose. The velocity acceleration in the nozzle results in a high velocity stream that is used to fight fires from a distance.
Turbulence: Turbulence occurs when a high velocity stream of water enters low or no velocity water. Vortexes form at the border region of the two bodies of water. The vortexes spin water backwards and perpendicular relative to the direction of the high velocity stream. This can be thought of as “nature’s mixer”.
A simple example of this is shooting a sharp stream of water into a bucket filled with standing water. Large amounts of turbulence are generated between the fast and no velocity water.
An example of both velocity acceleration and turbulence is an occluded blood vessel, as shown below. As blood, with its suspended cells, is squeezed through the occlusion, it undergoes velocity acceleration (from the equation above). As it enters the lower velocity blood past the occlusion, turbulence results. Even though blood vessels are soft and blood velocity is low, damage to cells from this process results in a higher risk of stroke.
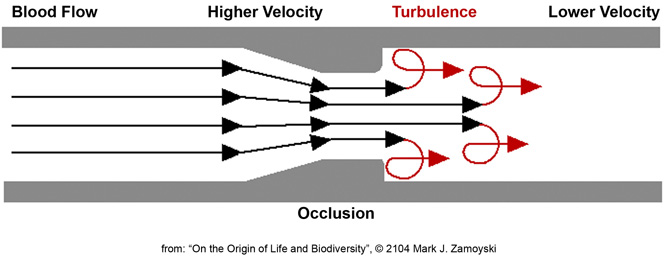
By boosting velocity and replacing the soft blood vessel with a hard skeleton labyrinth, we can begin to understand what happens in a CSFF. With the requisite fluid dynamics and molecular biology background we can now review the 5 selected CSFF structures to determine if they are capable of hosting the proposed grab bag DNA reassortment process.
Meet the CSFFs (Calcium Secreting Filter Feeders)
Photos of the external views of some of the fossilized structures built by the CSFFs are shown below:
 >
>
However, sectioned specimens are where the action is, as they provide the roadmap of how the labyrinth like structures channeled moving ocean water. Sectioned specimens of a few such specimens are show below.
 >
>
It is almost impossible to remove the structure from the matrix, and a more practical approach is to cut the specimen in half (section) to reveal the hard skeleton structure preserved inside:
The lighter parts are the encasing calcium matrix. The darker parts are the hard skeleton structure.
An artist’s illustration of what the specimen above, called the Permian Protopharetra, likely looked like, is shown below:
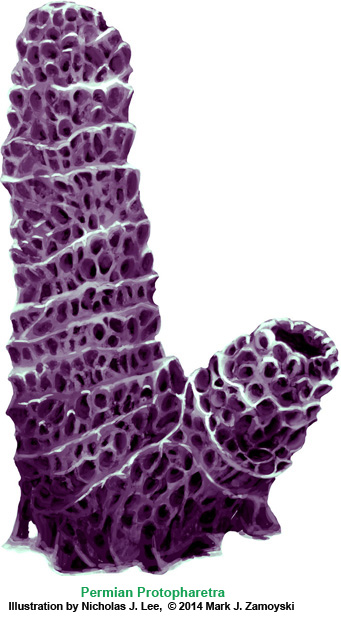
This specimen is morphologically identical to the Cambrian Protopharetra, an irregular archaeocyathan, (Boardman et. al., Fossil Invertebrates, 1987, p. 114 Fig. A and B), with the notable exception that the Permian specimen is about 5 times larger than its Cambrian counterpart. Archaeocyathans were the first unicellular eukaryotes that lived in colonies and secreted calcium carbonate as skeletal material. They were filter feeders, channeling moving ocean water through the hard skeleton labyrinth.
The Cambrian Protopharetra first appeared at the onset of the Cambrian explosion of multicellular life and then disappeared as the new multicellular ecosystem emerged. We have named the above specimen the Permian Protopharetra, which once again appears at the onset of the Permian multicellular explosion of life, and will once again disappear as the new multicellular ecosystem emerges.
Another example of a CSFF is the Spherical CSFF. The artist rendering is shown below:
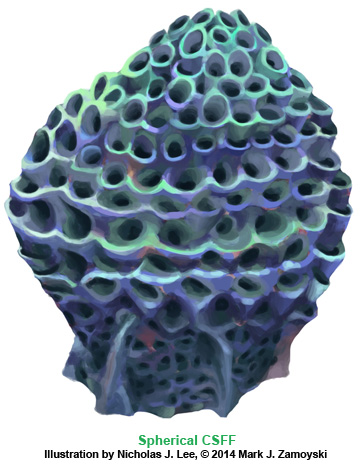
The sectioned specimen photo is shown below:
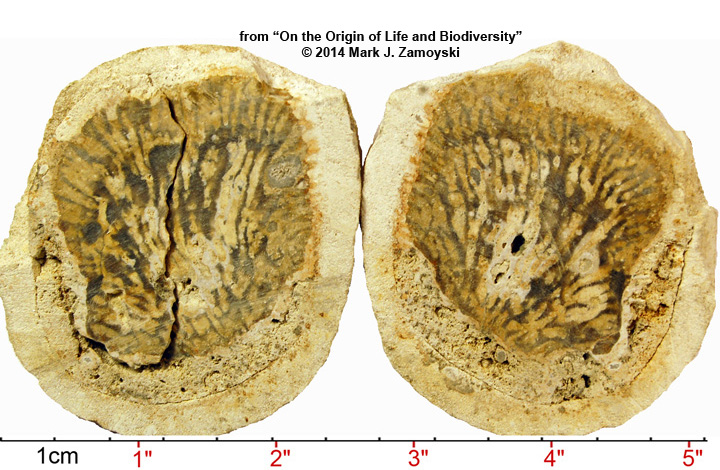
Examine the DNA Reassortment Structures
The sectioned specimens reveal internal structures that can be reviewed in context of fluid dynamics and molecular biology to see if they have the potential for host a DNA reassortment process that would account for both the transition from unicellular to multicellular life and also be capable of creating enormous biodiversity in the resulting life forms.
DNA Reassortment Structure 1: Nozzle and Slurry Chamber Structure
Starting with the spherical CSFF above, we can zoom in on one of the structures visible in the sectioned specimen on the left hand side. It is identified in the blue square:

The structure in the blue square above is further enlarged in two side by side pictures below:

The dark parts in the photo on the left outline the hard skeleton structure and the lumen is filled with the lighter colored calcium matrix . The photo (right) has the lumen / water channel in blue for clarity and depicts where the ocean water would have been when the CSFF was alive.
A tracing of the structure is shown below. Two noteworthy attributes are: 1) a nozzle structure that amplifies water velocity and 2) a post nozzle structure (slurry chamber) that enhances turbulence.
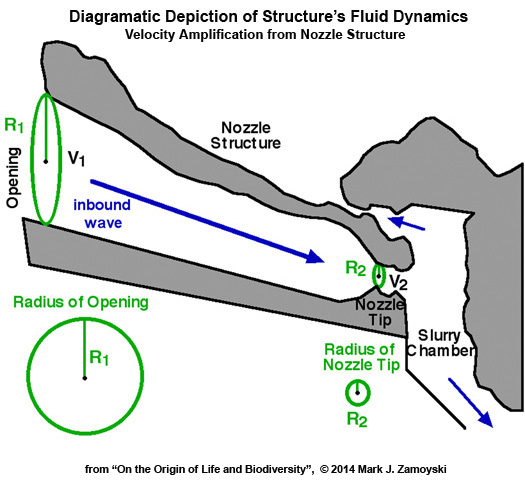
Water Velocity Amplification (~16X ): The opening on the ocean side of the nozzle orifice is more than 4 times larger than the nozzle tip that feeds the slurry chamber. For a given flow (e.g. in cubic mm / sec) coming in from the ocean, water velocity increases exponentially as the water passes through a confined space. The reason is that flow (Q) equals the velocity (V) times the cross sectional area (A) or Q= VA. The area (A) of a circle is A= ΠR2 where R is the radius and Π= 3.14. For a given Q, a reduction in radius results in an exponential reduction in area (i.e. R2), which in turn requires an exponential increase in velocity to maintain equality.
For a circular pipe: Q = ΠR2V or V = Q / ΠR2
For a given Q, velocity at the opening is V1 = Q / ΠR12 and velocity at the nozzle tip is V2 = Q / ΠR22
Accordingly, for a given Q, the velocity will increase exponentially with a reduction in radius. If the radius is reduced 4 fold, the velocity will increase 16 fold (i.e. 42).
The 16 fold velocity amplification shooting out of the nozzle tip into the perpendicular wall is analogous to accelerating a car from 10 mph to 160 mph by the time it hits the brick wall.
Turbulence. As previously discussed, turbulence occurs when a high velocity stream of water enters low or no velocity water. Vortexes form at the border region of the two bodies of water. The vortexes spin water backwards and perpendicular relative to the direction of the high velocity stream. This effectively functions as “nature’s mixer”.
Turbulence or Vortexes (shown in red) could be expected to form in several places as this high velocity stream enters and travels through the no or low velocity water in the slurry chamber. Some of these anticipated turbulence zones are shown below in red:
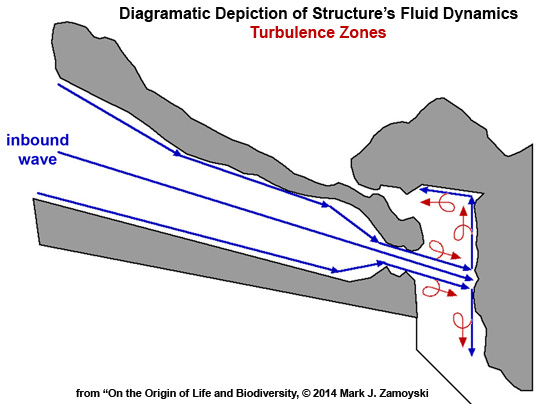
Combining fluid dynamics with molecular biology we can now review the structureπs mechanism of action to see if it has the means to achieve the proposed genetic reassortment.
 >
>
An inbound wave with its 1,000,000 suspended cells per ml is accelerated 16 fold and smashed into a perpendicular hard skeleton wall (back of the slurry chamber).
While the above is a highly simplified macro level view, at a micro level, the calcium carbonate "cement like" surface would look more like sandpaper. Cells moved anywhere along the way would be sheared open, like a grape being dragged across sandpaper. The spilled intracellular contents and sheared cell walls would also be moved into the slurry chamber.
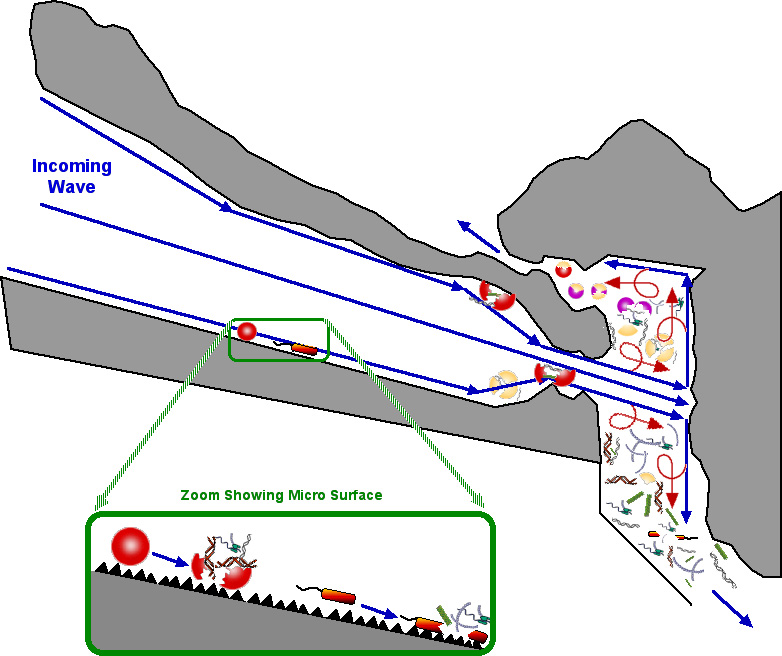 >
>
The ruptured cells in the slurry chamber are subjected to turbulence as the high velocity stream enters and travels through the low (or no) velocity water in the slurry chamber.
As the lighter lipid membranes spontaneously assemble, they take a gulp of the genetic slurry and can escape through the top vent. Heavier proteins and DNA fragments settle downwards, presumably toward the feeding colony.
The structure fulfills the requirements for hosting a grab bag DNA reassortment process.
While the intended purpose of the structure was apparently to shear open cell membranes to release the intracellular nutrients for the feeding colony, the unintended consequence appears to be a process capable of creating enormously biodiverse life forms from single celled organisms suspended in water.
This attribute is not unique to the spherical CSFF. The Permian Protopharetra specimens have numerous structures capable of hosting this type of DNA reassortment process. They contain at least 5 identified structures that can do this, with cyclonic based structures used extensively in their hard skeleton designs.
DNA Reassortment Structure 2: Cyclone Structure
The preferred structures used by the Permian Protopharetra class of CSFFs are cyclones, as well as advanced versions of supercharged cyclonic structures.
A good example of cyclones is found in "Mega Protopharetra", which appears to be a hybrid between a Permian Protopharetra and a Spherical CSFF. The dual cyclone structure would be able to utilize both inbound and outbound waves for a shred cycle.
A photo of the fossilized Mega Protopharetra specimen, along with an illustration, is provided below. The blue arrow highlights the area of the cyclone structures.
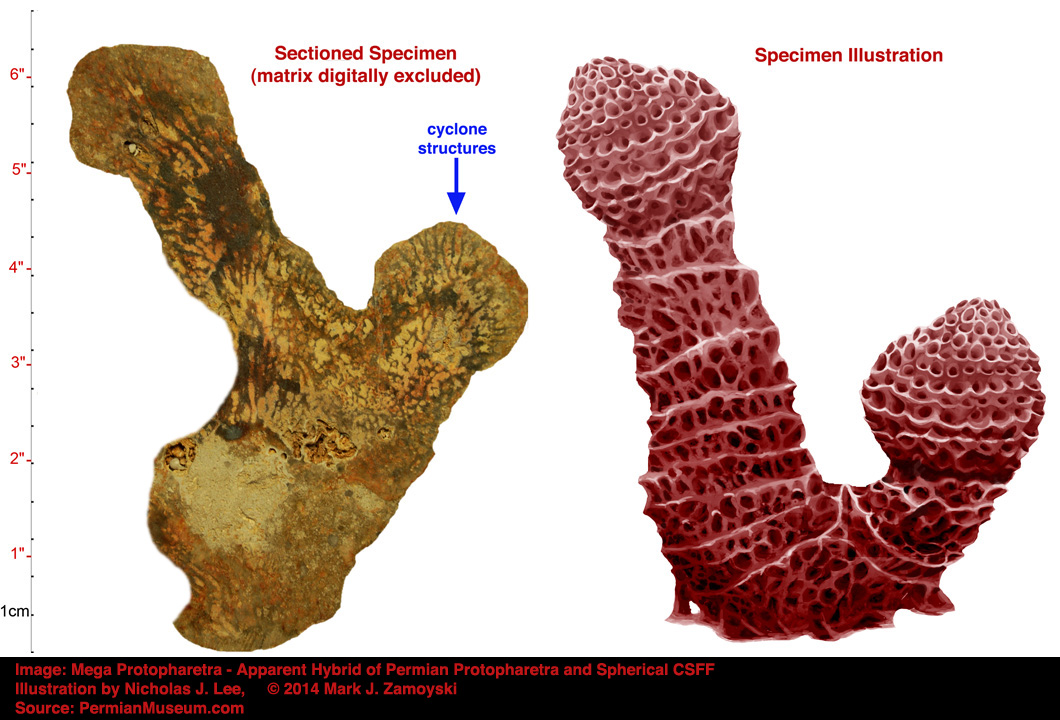
The photo below is a zoom of the fossilized structure, a view showing the isolated structures and water lumens, and a diagrammatic depiction of the functional cyclone structures.
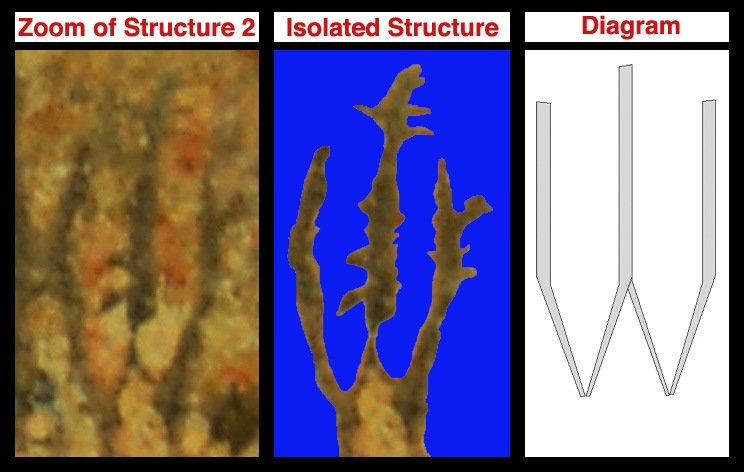
While the cyclone diagram shown is smooth for simplicity, it is important to note that at a cellular level the surface would be more akin to sandpaper. Pressing and dragging a cell across a limestone surface would be akin to dragging a grape over sandpaper. The actual depiction would more like the image below:
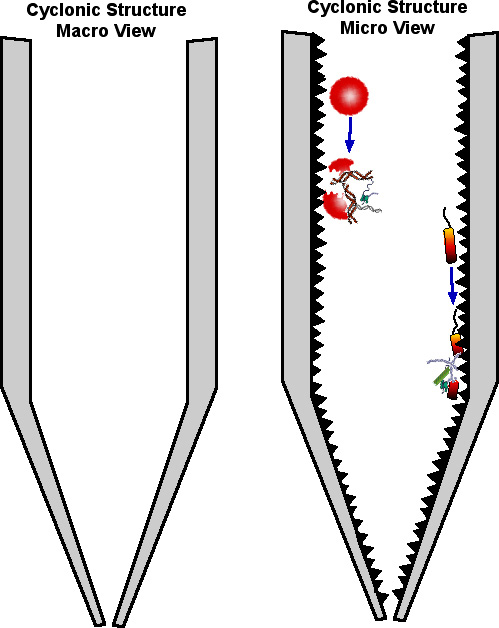
That having been said, we will proceed with the simplified macro view in the drawings below.
A simple cyclonic structure operates when a perpendicular stream of gas or fluid (water in this case) is introduced against the walls of a cylinder. This results in a stream that spirals down the wall, centrifuging larger suspended particles against the walls. As the cyclonic structure tapers, the area is reduced and the water velocity exponentially increases (per the velocity and area equations previously discussed). As the stream reaches the bottom of the cyclone, it begins to flow upward through the center of the spiral and exits the cyclonic structure.
While a cyclone is a passive structure, the shallow ocean provides the action in a 4 stroke cycle of 1) Incoming Wave, 2) Calm Water, 3) Outgoing Wave, 4) Calm water. The combination of the 4 stroke cycle of the motion of the water (containing 1 million suspended cells per ml ) with the dual cyclone structure is depicted diagrammatically below:
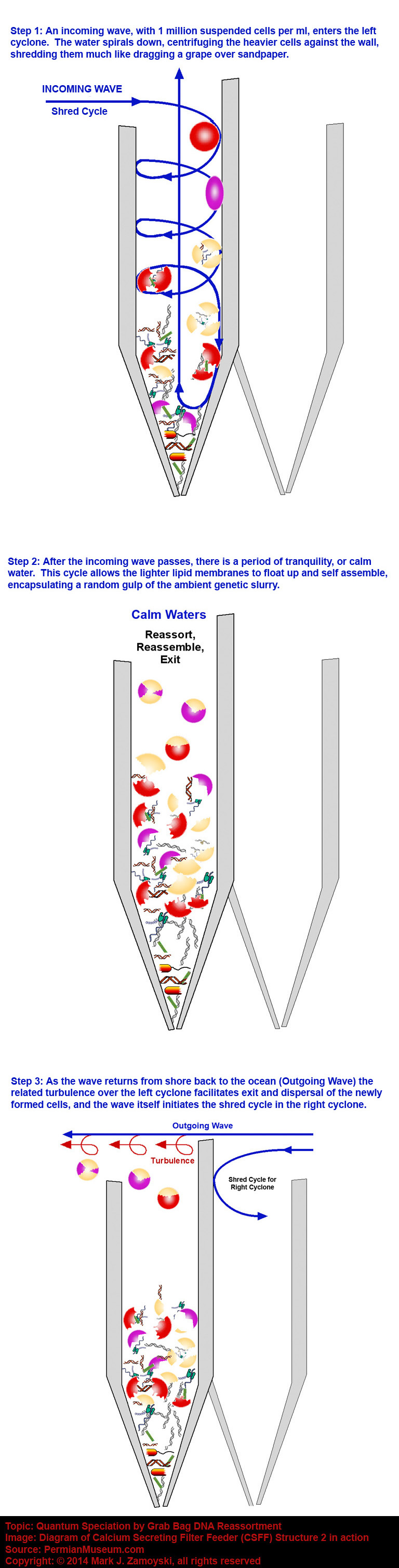
A cyclone structure also fulfills the requirements for hosting a grab bag DNA reassortment process.
While the first edition of "On the Origin of Life and Biodiversity" covered several other CSFF structures capable of hosting grab bag DNA reassortment, author has decided not to labor the point in this second edition, but instead present the potential for natural geological structures to accomplish the same thing.
CSFFs have highly specialized structures for shredding and reassorting cells, and their appearance in the fossil record during the Cambrian Explosion of Life and Permian Explosion of Life is not likely a coincidence.
However, It should be noted that the hosting of DNA reassortment may not be exclusive to CSFFs.
Rocks containing cavitation, when combined with the motion of waves, could also serve as genetic reassortment machines. Internal chambers, as they filled with shredded cell contents, could also serve as incubators for the new, reassorted life forms.
As an example, progressive sectioning of a rock with such geological cavitation reveals the complexity of multiple levels of shred surfaces as well as various internal cavitation chambers.
Progressive Sectioning of Rock with Geological Cavitation

Naturally occurring geological cavitation, combined with moving water, and functioning as shredders / reassorters, likely functioned at a much lower level to create new life forms long before the CSFFs appeared and long after they disappeared once a new ecosystem was established. Cavitation, combined with moving water, still likely functions to this day, and the creation of new life in the ocean is not likely a rarity, but the survival of a new life form to maturity in an established ecosystem would be the rarity.
The explosion of unicellular eukaryotic life 1.5 Ba, with its single lipid bilayer cell wall and mitochondrial DNA that was a reassortment of DNA from several types of bacteria (prokaryotic cells), may have been hosted in naturally occurring geological structures. Among the new life forms created were the single celled eukaryotic organisms (archaeocyathans) that built the CSFF structures, to obtain nutrients from cells suspended in moving water, with the unintended consequence of hosting genetic reassortment and a resulting "Explosion of Life".
It is now time to take a look at the empirical data (i.e. fossil record) of the new life forms spawned by this genetic reassortment process during the "Permian Explosion of Life".
Chapter 4
Fossil Record of Life Forms Spawned by DNA Reassortment
The output of the genetic reassortment reassortment process is dependent on the input available for that process. Therefore, it is essential to understand both the pre-existing unicellular life forms, as well as the multicellular extinction survivors, as both served as input into the reassortment process.
The Input
Unicellular Prokaryotic and Eukaryotic Organisms
The first, and perhaps most important, input into the process are single celled prokaryotes and eukaryotes. Unicellular prokaryotes first appeared some 3.8 Ba and the unicellular eukaryotes appeared some 1.5 Ba. In terms of biomass, they are the most prevalent life form on the planet today. One gram of soil contains 100 million to 1 billion bacterial cells. Coastal oceans contain 1,000,000 cells per ml.
Mitochondrial DNA: Mitochondria is a cell’s power plant that stores energy from aerobic respiration (metabolism of glucose). Mitochondria is of prokaryotic origin: its DNA is separate from that of the nucleus, is circular (bacterial), and its nucleotide sequence analysis points back to early bacterial origins (rickettsia, rhizobacteria, and agrobacteria (Alberts et. al., Molecular biology of the Cell, Third Edition 1994). Physiological life can be defined as a collection of concentration gradients. The presence of concentration gradients means life. The absence of concentration gradients means death. It takes energy to maintain concentration gradients. Without the prokaryotic mitochondrial DNA that arrived on earth some 3.8 Ba, life as we know it today would not exist.
Cilia and Flagella DNA: Unicellular organisms can use flagella, cilia, or pseudopods for motility. This ancient DNA made its way to present day humans. In humans, the sperm cell uses a flagellum to propel itself. Cilia drive the movement of the mucus blanket that sweeps dirt out of the lungs. Beating of cilia in the fallopian tubes moves the egg from the ovary to the uterus. Pseudopod DNA likely went on to form limbs for mobility.
Chemotaxis DNA: Chemotaxis is the initiation of motility (activation of flagella, cilia, or pseudopods) in response to chemicals in the environment and is used by unicellular prokaryotic and eukaryotic cells to find and move toward food. In humans, cells such as neutrophils, the body’s first line of defense against bacteria, recognize chemicals produced by bacteria and move directly toward them. Additionally, tissue resident mast cells are activated by antigens and in response release chemotactic factors such as Eosinophil Chemotactic Factor A, Chemotactic Factor (NCF IL8) and Leukotriene B4, which in turn result in chemotaxis of a broader set of immune system cells toward the site.
Cell Signaling DNA: Unicellular organisms have cell signaling capability in colony situations that causes the outermost cells to differentiate and form a hardened protective outer layer. Skin may hail in part from this DNA. Skin is an extremely important feature for multicellular life, primarily because of its ability to contain an aqueous environment which allows atoms to exist as ions (Na+, Cl-, K+, Ca++) which in turn allows for maintenance of concentration gradients including electrochemical gradients. A 70 kg human (~ 70 liters volume) is made up of ~ 10 liters of cells (~ 10 trillion cells) bathed in 40 liters of extracellular fluid, with the balance made up of bone, fat, muscle fibers and connective tissue. Everything is contained within a skin sack. The resident extracellular saline solution is our gulp of the ocean we needed to take before we could step onto land. Skin also revolutionized cell signaling. Cell signaling is the production of chemicals (e.g. testosterone, estrogen) by a cell that alters DNA expression of distant cells. In a closed environment, the chemical signals are not washed away by the ocean, but can more effectively reach their intended target cells.
Skeletal Matrix DNA: Archaeocyathans are the first known unicellular eukaryotes that secreted calcium carbonate as a skeletal matrix ( i.e. the Calcium Secreting Filter Feeders). They first appeared some 500 million years ago and also built the hard skeleton labyrinth structures that hosted the DNA reassortment process during the Permian period. Their DNA may also have found its way into an osteoblast cell, which is the bone building cell in boned multicellular life forms.
The point is that a significant part of multicellular eukaryotic DNA traces back to ancient unicellular origins.
Extinction Survivors - Multicellular Eukaryotic
While 96% of marine life was wiped out in the last Permian extinction pulse, that means 4% survived and were available as input for the genetic reassortment process. While the Cambrian explosion of life had unicellular life as an input, the presence of multicellular life during the Permian explosion would have stacked the deck for a much more robust explosion of life to occur. The survivors would also stack the deck toward "their form" of life form.
Each cell from a multicellular life form contains the entire genome, or the DNA that codes for all of the cell types made by that multicellular life form. As an example, the human genome codes for more than 200 different cell types, and each cell contains the DNA for all of the different cell types. A cell from a multicellular life form can enter the ambient environment in numerous ways such as skin shedding, injury, death, excretion of body fluids, etc..., much in the same way cells are obtained for DNA evaluation in criminal cases, including contact DNA testing. Any such liberated cell can become input in the genetic reassortment process, with the entire genome being available for reassortment, or parts thereof.
Nematodes: Digestive, Reproductive, Muscle, Nerve, and Skin DNA
Nematodes (roundworms and eelworms) are the oldest known multicellular life forms and their explosion of life was almost as spectacular as that of the unicellular prokaryotes. Today, they represent 90% of all animals on the ocean floor and often exceed a million individuals per square meter in earth's topsoil. There are over 25,000 known species, ~ 35 species are known to infect humans, and some scientists estimate as many as half a million are yet to be discovered.
CDC Photo of Intestinal Nematodes
Nematode related fossils found date back to the Cambrian period (543-490 Ma) but because many nematodes are microscopic, and others are transparent, their discovery as fossils is difficult, and some estimates put their origins back to Pre-Cambrian times. Nematodes are among the multicellular survivors that made it into our fossil collection.
Nematodes have a tubular digestive system, with a mouth at the front and anus at the back. Nematodes have internal male, female, or hermaphroditic reproductive organs, where eggs are produced and pass out the digestive tract into the environment. They are invertebrates that have an exoskeleton made up of a hard, but flexible, cuticle on the outside (secreted by their epidermis). Nematodes have 2 nerves, one on the back and one on the belly, that run the length of the nematode. The nerves connect to muscles attached to the cuticle, allowing for side to side (fish like) motion.
Nematodes are basically a motile intestine, with a mouth, anus, and gonads (glands capable of producing haploid cells). Nematodes were effectively the core blueprint for all multicellular life forms that followed, all the way through to humans.
Because unicellular prokaryotes and eukaryotes, multi-cellular nematodes, and CSFFs were present during both the Cambrian and Permian Explosions of Life, a subset of our fossils should provide a rare glimpse as to what some of the earliest DNA re-assortments looked like during the Cambrian explosion of life. This subset shows how nematode DNA developed into boneless fish in both periods, and then with the integration of CSFF DNA, went on to become boned fish. Boned fish like the lung fish in turn are believed to have been the forerunners of terrestrial tetrapods.
Unexpectedly, fossils in the output section below provide evidence that the "uprights", both unboned and boned, may have also come from nematode origins.
Nematode nerves were highly conserved through time and the same neuron related proteins in transparent roundworms such as Caenorhabditis elegans are also found in humans today. These nematodes are used today as models to study the proteins produced in response to nerve injury and healing, in order to better address nerve injury and degenerative diseases in humans.
Nematodes have most of the DNA for many of the life forms that followed, with the exception of bone DNA, which appears to have come from the Calcium Secreting Filter Feeders (CSFFs), as we will see in the output specimens. The addition of bone to soft tissue was an enormous leap in multicellular life form capabilities. It was not just the structural function of a skeleton that allowed for mobility rather than just motility, but bone is at the core of many processes crucial for multicellular life.
Bone reservoirs and releases growth factors, phosphorous, and calcium in response to chemical signals. The released compounds are used for numerous processes including cell population density management and injury management, energy storage, activation of proteins, depolarization of nerve membranes, enhanced neurotransmitter release, enhanced muscle function, and propagation of the calcium wave that underlies consciousness and memory formation. All of these are covered in detail in Chapter 5.
The external view of some fossilized nematodes on ocean floor, encased in the typical limestone matrix, as well as a sectioned specimen are shown below. The sectioned specimen reveals a transparent nematode, where the only truly distinguishable features are the mouth and "eye dot" at the far left part of the life form:
Nematodes on Ocean Floor and Sectioned, Transparent Nematode

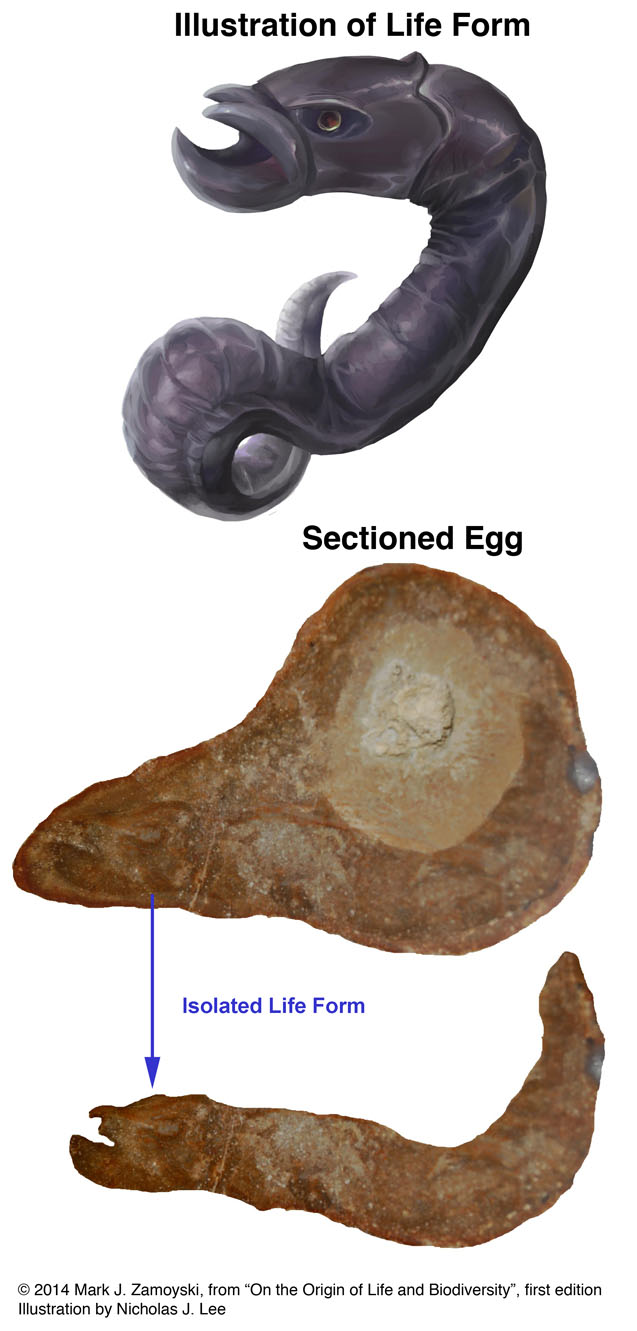
A sectioned egg showing an almost mature, slightly more distinguishable "jawed" nematode is shown below:
Almost Mature "Snake Like" Nematode in Sectioned Egg
Lung Fish: Lung DNA, Tetrapod DNA: As for the more advanced fossilized aquatic survivors we have found to date, perhaps the most significant is the Lung Fish.
Arizona Permian Lung Fish

Lung fish first appeared some 400 million years ago (during the explosion of fish life) and three genera of lung fish are still alive today, one on each of three continents (Australia, South America, and Africa). Lung fish are extreme survivors. They can use their pseudopods to move on land. They use aestivation (dormancy) burrows when their pools dry up. They breathe air through their swim bladders and drop their metabolic rate. Lung fish aestivation burrows are found in the fossil record during the Devonian (~ 400 Ma), Carboniferous (~ 300 Ma ), and Permian (~ 250 Ma) periods.
Lung fish are believed to be forerunners of terrestrial tetrapods (four legged air breathers that walk the earth). Their lung is a modified swim bladder that can absorb oxygen and remove waste. Their 4 limbs are in the same relative position as those of terrestrial tetrapods.
Shark: Advanced Eye DNA:Another significant fish that made it into the collection is the shark. Sharks are cartilaginous fish that date back to more than 420 million years ago and are found in all seas today. Shark eyes are similar to the eyes of vertebrates and include similar lenses, corneas, and retinas.
Arizona Shark
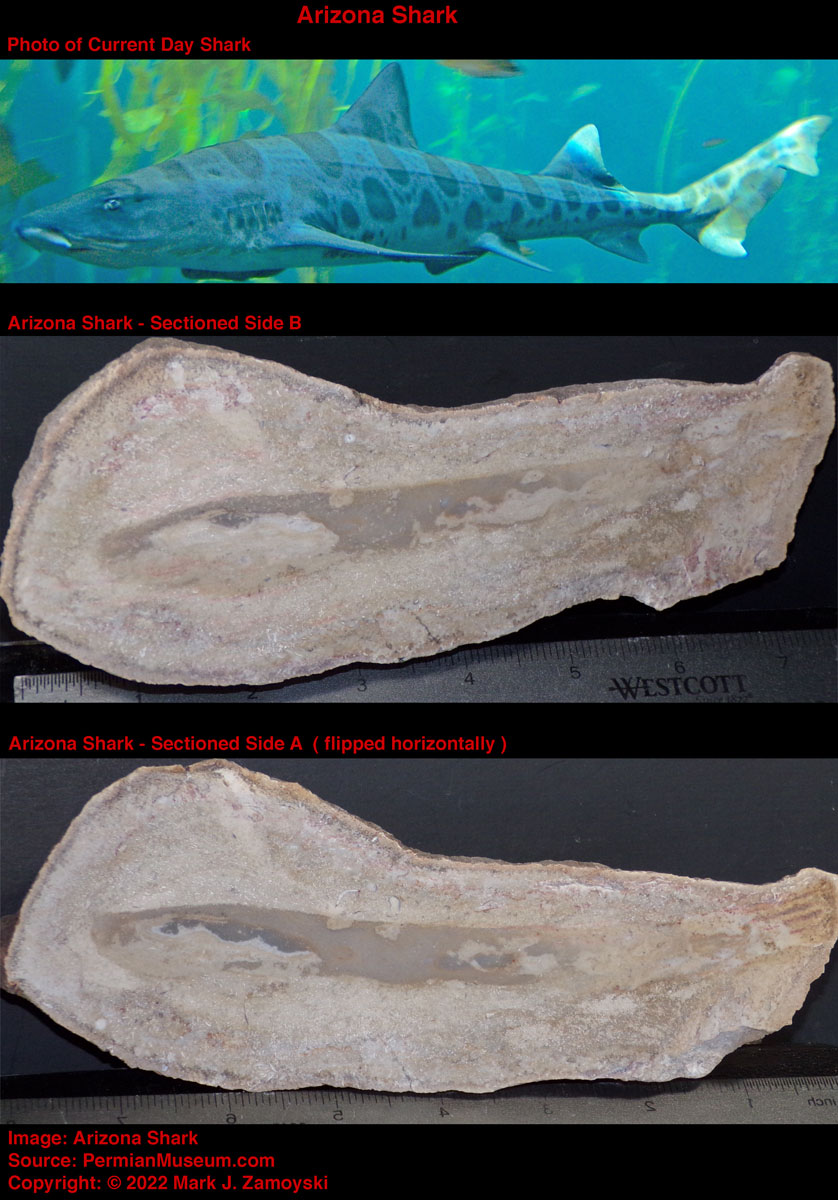
Accordingly, the DNA for an advanced ocular system was available for the genetic reassortment process, and it is the dominant form of eye in use today.
Sting Ray: Wing DNA:Another significant survivor that made it into the collection is a stingray, in part because of its "wing" DNA.
Arizona Stingray - Upper Matrix Removed

Stingrays first appear in the fossil record some 450 Ma. They are cartilaginous fish with pectoral fins or "water wings" for propulsion. There are more than 200 species of stingrays alive today. Sting rays are related to sharks.
Yet another survivor that made it into the collection is the jellyfish. Jellyfish are gelatinous umbrella of bell shaped life forms that pulsate, or use contractile motion, to achieve propulsion for locomotion. Jellyfish have been in existence for at least 500 million years. Jellyfish survived the Permian period and are found worldwide today. Their fossils appear mostly as traces that are part of the limestone. As an example:
Fossilized Jellyfish

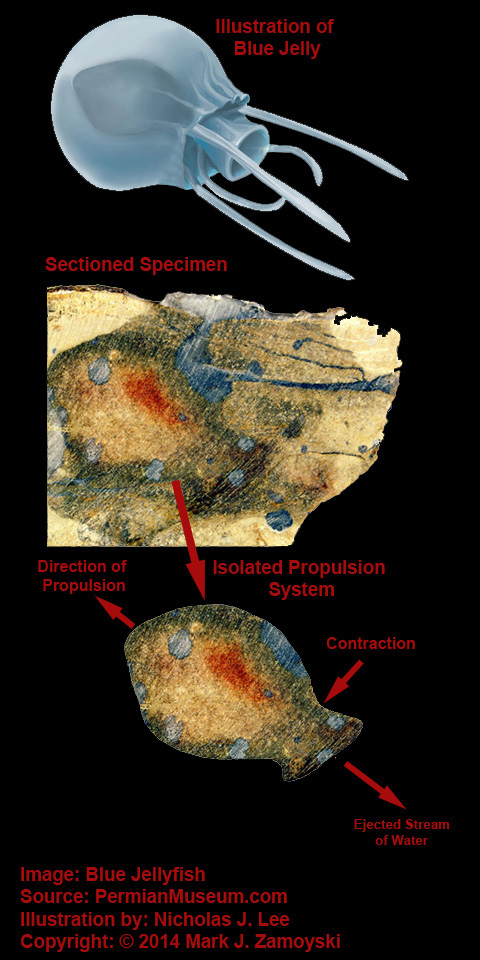
As another example, the specimen we call Blue Jelly has a internal contractile sack that is visible:
Blue Jelly
Reptile DNA Reptiles first appeared around 312 Ma. The DNA of mammals and reptiles is 89% the same.
Reptile on a Rock

While reptilian DNA lives on in lizards, crocodiles, snakes, worm lizards, and turtles, its contribution to mammalian life forms is not trivial. The word dinosaur is literally from the Greek "deinos" for terrible and "sauros" for lizard.
In addition to the whole body fossils of aquatic life forms presented above, there were also partial remains found that included what appear to be bone from multicellular survivors, including bones, tusks or fangs, and pieces of skin. Their presence is significant in that the more advanced DNA would have been available as input into the reassortment process, allowing for more advanced life forms than those that emerged from the Cambrian explosion of life.
Some examples of these "pieces" are shown below:

Iron rock impacted into limestone was found in proximity to these pieces, raising the possibility of a high pressure wave that blew these life forms apart. It is not clear if this is related to the major Permian extinction events or an unrelated localized extinction event.
Iron Rock Impact Near "Pieces"

Now that we have finished an overview of the input, we can next take look at the output.
Output Specimens
The grab bag DNA reassortment process previously disclosed could be expected to generate several types of new life forms:
1) Life forms that appear to be quantum leaps backward in evolution could be expected, as primitive unicellular DNA was reassorted and recombined, in what would be virtually identical to what happened during the Cambrian Explosion of life. While these types of life forms were new 500 Ma, they would appear to be a quantum (250 million year) leap backward in evolution when viewed from a Permian perspective.
2) Biotic Hybrids and DNA Amalgamations would also be expected. Amalgamations is used in context of central DNA in a life form representing features from various life forms. DNA Amalgamation combinations such as fish / reptile life forms, bird / fish life forms, and bird / insects life forms, could be expected, as DNA from very different life forms could combine into new, viable life forms.
Biotic Hybrids is used in context of two life forms, with separate DNA, living biotically as a single organism. A contemporary example of a biotic hybrid is lichen, which is a composite organism made up of algae or cyanobacteria living among filaments of various species of fungi. Lichens have different properties from those of their component organisms.
New life forms could also be both DNA Amalgamations and Biotic Hybrids. A contemporary example of such a life form is a human. A human is an amalgamation of 46 strands of DNA, coupled as 23 pairs, contained in the nucleus of every cell. The notable exception is mitochondrial DNA, which is circular, of bacterial origin, and exists outside of the nucleus in the mitochondria, which could arguably be classified as a biotic hybrid. When you then include the thousands of different types of bacteria living in our gut and on our skin, we are unquestionably also biotic hybrids.
3) Duplicate structures, and duplicate life forms living as one, could also be expected. Redundancy is not prohibited but expected, but would only be seen in the fossil record if the resulting life form was viable.
4) Radical Amalgamations: Incorporation of extremely unrelated DNA, into multicellular life forms, could also be expected. The results could radically change the direction of life on earth. As an example, the integration of CSFF DNA into soft tissue life forms could spawn the emergence of boned life forms. The integration of CSFF DNA with other life forms would be highly expected because of the proximity of CSFF DNA to the reassortment process, and hence its availability for inclusion in the reassortment process.
5) Future Life Form Forerunners could also be expected. After a mass extinction event, in the absence of established predators, the stage would be set for a new ecosystem to arise. Some of the newly created life forms could be expected to become part of the new ecosystem. Either forerunners, or the new life forms themselves, should be somewhere in the fossil record, starting from the reassortment period(s), through the subsequent re-speciation period, and possibly into the new stable ecosystem period.
6) Unrecognizable or "indeterminate" life forms would also be expected. Only a small percentage of what was generated by DNA reassortment could be expected to be viable, and even a smaller percentage would find its way into earth’s play book of life. Some life forms could be so different that we would not have a frame of reference to compare them to.
All of the above are actually observed in our fossils, and are presented in the sections below.
Fish and Fish / Lizard Combinations
Fish first appeared some 500 Ma, and the Fish Explosion of Life occurred around 400 Ma. Reptiles first appeared around 312 Ma. There are many normal looking fish in our collection, however we have not yet had the time or expertise to identify which of them are pre-existing survivors and which are new.
The fish presented below are the ones that are more suggestive of being the product of the genetic reassortment process.
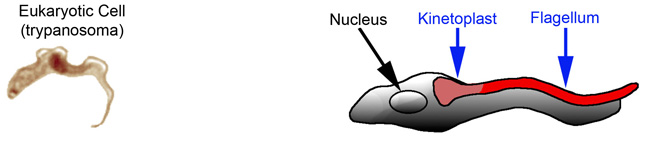
Prokaryotic cells can only use a rear mount flagellum for propulsion because of their rigid, triple layer cell wall.

Eukaryotic cells have a flexible lipid bilayer cell wall, allowing them to use either a rear mount flagellum or side mount flagellum. An example of a side mount flagellum is in the unicellular trypanosoma, which causes sleeping sickness, and is only about 25 µm long.
One example of an early life forms generated by the genetic reassortment process appears to be the flagellate unicellular giant shown below, which has a rear mount flagellum. At 2 inches long, it is 2,000 times larger than the unicellular trypanosoma. The observable flagellum appears to be the same as that coded for by DNA of unicellular prokaryotes.

Flagellate Unicellular giant was likely similar to something that would have been spawned during the Cambrian explosion of life, when only unicellular organisms were available for input into the genetic reassortment machines. It was motile in water and 2" long so we have included it in the fish section.
The Flagellate Unicellular giant is an example of an expected "Quantum Leap Backward" life form.
A slightly more advanced, yet still "Quantum Leap Backward" life form is Flagella Fish shown below, which has a side mount flagellum instead of a fish tail. At 5 inches long, it is 5,000 times larger than the unicellular eukaryotic trypanosoma. The observable flagellum appears to be typical of the unicellular eukaryotic DNA that codes for a side mount flagellum on a flexible bodied life form. However, the two arm like paws at the front indicate the possibility of more advanced bilateral, pseudopod DNA.

Some of the early fish are morphologically similar to nematodes. They appear to be "super-sized" nematodes with a more pronounced fish like head and early fish tails.
Nematode Fish 1
It also appears that some nematode fish integrated CSFF DNA. This appears to form an internal "skeletal matrix" onto which muscles could attach, in addition to or instead of, the exoskeletal cuticle attachments. CSFF's are single celled organisms that secrete calcium onto a skeletal matrix. They may have been forerunners of Osteoblast cells (bone builders) in boned life forms. We see this integration of CSFF DNA in several other life forms.
One of our specimens (Indeterminate 22) is morphologically similar to the boneless nematode above, but has CSFF DNA integrated internally. This may have been a forerunner of boned aquatic life forms. It also serves as a representative example of how other life forms may have became "boned life forms". It also appears to have more advanced eye DNA than nematodes.
Nematode Fish with CSFF DNA Integrated

Morphologically similar life forms to the two above may have also been of nematode origin, but with an "upright" orientation (i.e. vertical or "upright" orientation of body, with head perpendicular to body). The specimen below shows one such "upright" life form, without bones, but with a more advanced eye.
Upright 1, Boneless but with Advanced Eye DNA
The second specimen is an upright, but with visible CSFF DNA integration, forming a skeletal matrix.
Upright 2, CSFF DNA Integrated

Getting back to fish, another example of a "Quantum Leap Backward" life form is the Jawless Fish shown below:

Jawless fish first appeared in the Cambrian Period, but were believed to have gone extinct during the Devonian period (420 - 360 Ma). It is not surprising that fish like the Jawless Fish would go extinct, as they are defenseless, small, and instantly relegated to the bottom of the food chain when meaningful predatory fish arise. While their reappearance during the Permian quantum speciation by grab bag DNA reassortment would be expected, their tenure could also be expected to be much shorter because of the established presence of advanced predatory fish at the time.
A possibility is that the inclusion of the DNA that coded for these types of life forms, in the original prokaryotic cells that arrived some 3.8 Ba, was to provide an easy food source for emerging multicellular life forms (i.e. small, defenseless life forms would simply have been "manna of the ocean").
An example of a "Amalgamation / Combinational Life Form" is the specimen below, that appears to have the mouth and body of a stone fish, but a contractile sack propulsion system of a jellyfish. It is not a normal fish as we know it, and as such may be a genetic reassortment product of the 2 different life forms, which resulted in a combination that was still viable.
Stone Fish Hybrid - Sectioned Specimen
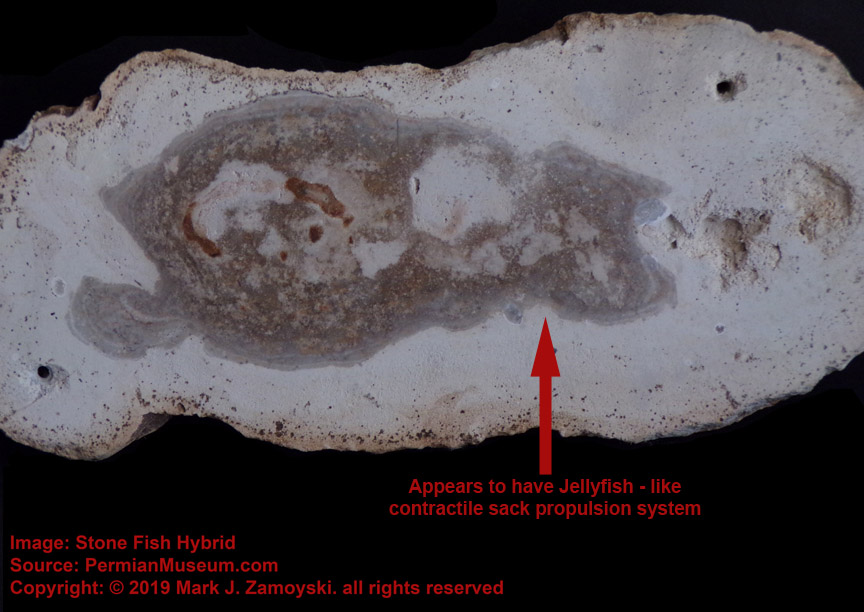
Moving to the category of "Future Life Form Forerunners" by genetic reassortment is the specimen called Whaleen, which resembles a truncated Baleen Whale in existence today.
A photo of the external view (Side A) of Whaleen is shown below. The specimen is ~ 5 inches or 12 cm long.
A photo of the sectioned specimen ( Side B) reveals a fossilized optic neuron in the upper left of the sectioned specimen. A cutout of the optic neuron is included above the actual neuron for clarity. The optic neuron is connected directly to a motility group, and the organism has no brain.
The example above is relevant because a neuron is a separate cell type. A neuron is not the product of an adaptive evolutionary improvement to an existing structure, but the introduction and integration of completely new DNA, as would be expected under genetic reassortment.
An optic neuron without a brain would likely result in mindless, relentless movement toward light, similar to how a moth repeatedly hits a porch light. If fish that existed prior to the Permian period had a brain, Whaleen would appear to be a quantum leap backward in evolution. However, it is exactly what would be expected by "starting over" from unicellular life forms.
In the ocean, moving toward light means moving toward food, as that is where photosynthesis based organisms (algae) thrive, as well as the related food chain that thrives on them. Even single celled organism (without an optic neuron) have the ability to orient themselves toward light, so this is a primordial trait.
The central lumen appears to have gestating progeny (preserved as blue translucent crystals). Reproduction inside the intestine is typical of nematode DNA. It is possible that the earliest womb was simply a sack encasing the progeny that allowed nutrients from the food lumen to reach the progeny, as well as protect the progeny from being digested. Internal birth, versus eggs that are subject to predation, is a trait that has survived to this day.
The neuron in Whaleen is fairly advanced, and may have come from an advanced Permian life form. It appears there may also have been more primitive optic neuron cells in use, as can be seen in the sectioned specimen we call "Sea Pig".
Simpler Optic Neuron
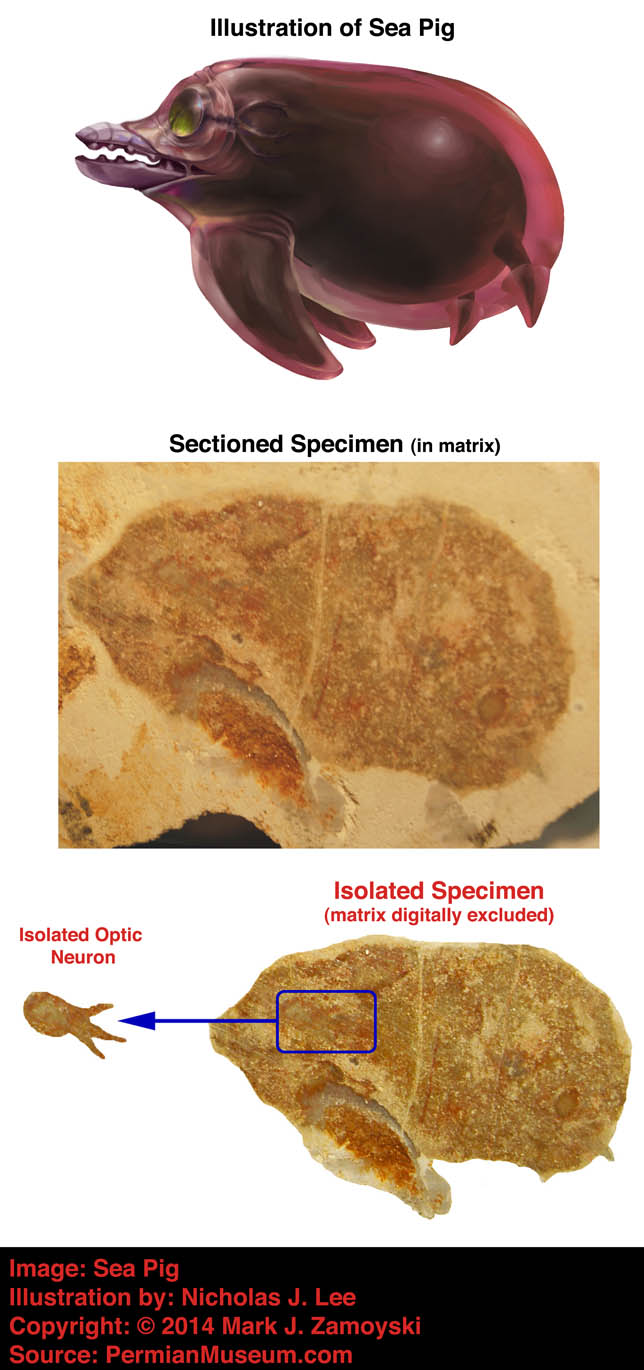
It appears one of the early integrations of bone cell DNA may have been simply as a defensive needle like bone structure protruding upward from the rear of a fish, as shown below:
Bone as Backside Defense
That single passive integration would have protected the fish from being taken by a backside attack. Since eyes look forward, an attack from the rear is often used by predators. The undigested stomach contents of the Zamoyski dragon, covered later in this chapter, show a fish swallowed whole, taken by an attack from the rear.
Defensive venomous spikes are still in use today in the Lionfish (Pterois) and non venomous spikes are used by the sea urchin (old French for hedgehog). The advantage in the more contemporary specimens is that the spikes surround the entire life form, offering protection from multiple directions, as shown below:
Lionfish (lower right) and Sea Urchin (upper left)

Another example of early integration of bone DNA may be "Tusk Fish" shown below, where the bone is embedded in soft tissue at the front of the fish.
Bone as Tusk
On the other side of the spectrum, there is also the potential for "over integration".
An example in the "Duplicate Structures and Duplicate Life Forms Living as One" category is Double Fish shown below. The genetic reassortment process does not necessarily have to generate a singular, clean life form. Life forms with redundant DNA, which may or may not be synergistic or even compatible, could be expected from the grab bag process. Many would not be viable, but a few could go on to maturity if the additional DNA was beneficial. Double Fish appears to be one such example. It appears to have both a pincer mouth on the exterior and a jawed mouth further back. While most specimens have either a linear central food lumen or a U-shaped lumen, Double Fish has both, indicating the possibility of two entire life forms in one.

Life forms with redundant or duplicate structures would be expected under the grab bag DNA reassortment process. The lack of boned structures likely allowed the more amorphous "body sack" to accommodate other structures, or even two life forms. The presence of both mouths directly in the food intake stream could have made two life forms viable, alternatively, the double mouth structure could also be viable in a single organism.
As an example of the latter, moray eels in existence today have a pharyngeal_jaw , or a second set of jaws, that reside in the throat. The arrangement is basically the same as that observed in Double Fish above.
The evidence of redundant structures are not unique to our fossil. An example of what appears to be a redundant upper jaw and nostril can be seen in the skull photo of Proterosuchidae which is found in the fossil record from 252 - 247 Ma. That double structure appears to add no value, and was likely more of an impediment to capturing prey, which is likely why that life form is no longer with us.
Genetic reassortment could also be expected to combine and integrate DNA from unrelated life forms ( e.g. reptiles and fish, birds and fish, birds and reptiles, etc... )
An example of a Fish / Reptile "Amalgamation / Combinational Life Form" is the Zamoyski Dragon specimen below. It has hard reptilian like skin but the body of a fish.

Sectioning reveals the life form was closer to a unicellular protist, as it has no bones or circulatory system yet. The stomach contents reveal a fairly intact undigested fish, implying Zamoyski Dragon used a typical protist approach of swallow whole and digest. The undigested fish has been named “Goby Shark” because it has has features of both a goby fish and a shark.
The mouth of the Zamoyski Dragon was likely akin to the tentacled mouth of a squid, shown below:

Specialized cell types would have been responsible for the hardened segmented skin around the head of the Zamoyski Dragon as well as for the backward facing barbs at the top rear, inside of the tail, making this a multicellular life form. The backward facing barbs at the rear would likely lodge in a pursuer’s throat, preventing swallowing and facilitating forward escape. They would be effective until a kill and chew world arose.
That segues us into the next specimen, which is part of the kill and chew world.
Another example of a Fish / Reptile "Amalgamation / Combinational Life Form" is Lizard Fish, which is similar to Zamoyski Dragon in respects, however it has a toothed jaw and head shape more akin to that of a lizard.
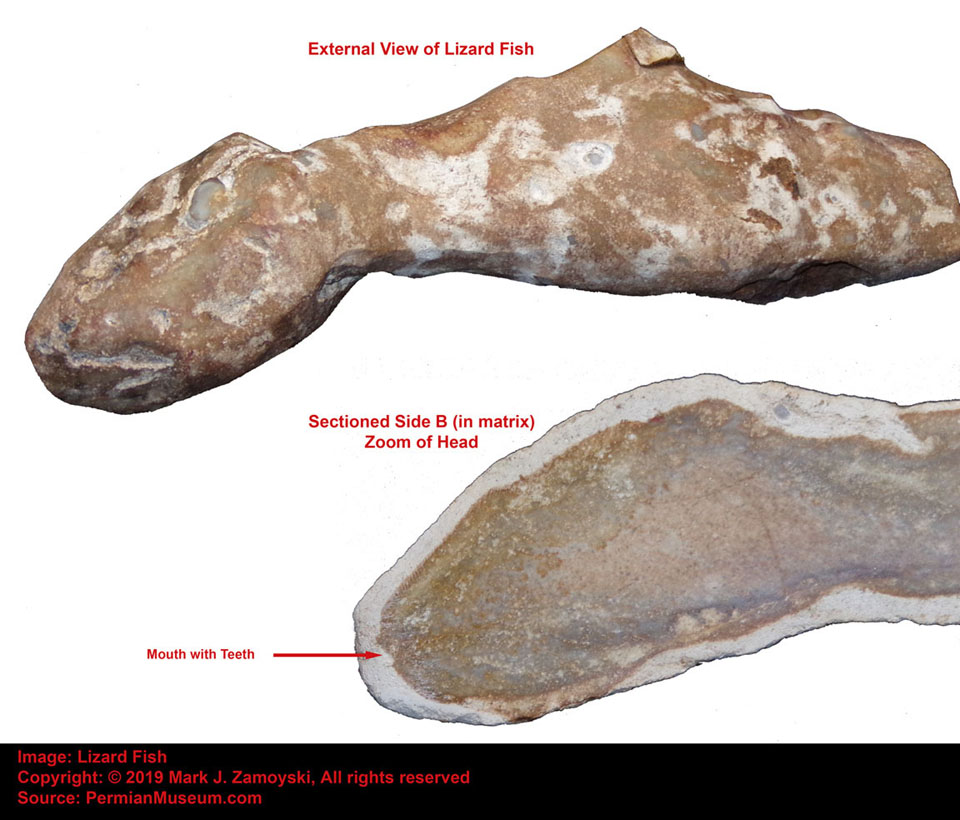
In addition to the toothed jaw, Lizard Fish has micro spikes on the outside of its skin. The small spikes on Lizard Fish are shown in the photo below, which is a zoom of the top of the head of the sectioned specimen in matrix.

Most lizards today are tetrapods (such as the "Reptile on a Rock" Fossil shown in the input section). Lizards run with a strong side-to-side motion, perhaps a vestige of their combined fish / reptile DNA (i.e. the fishtail motion).
Another aquatic or fish / lizard "Amalgamation / Combinational Life Form" called Geckosaurus is shown below. It has the head of a dinosaur such as T-Rex and the body of an aquatic life form.
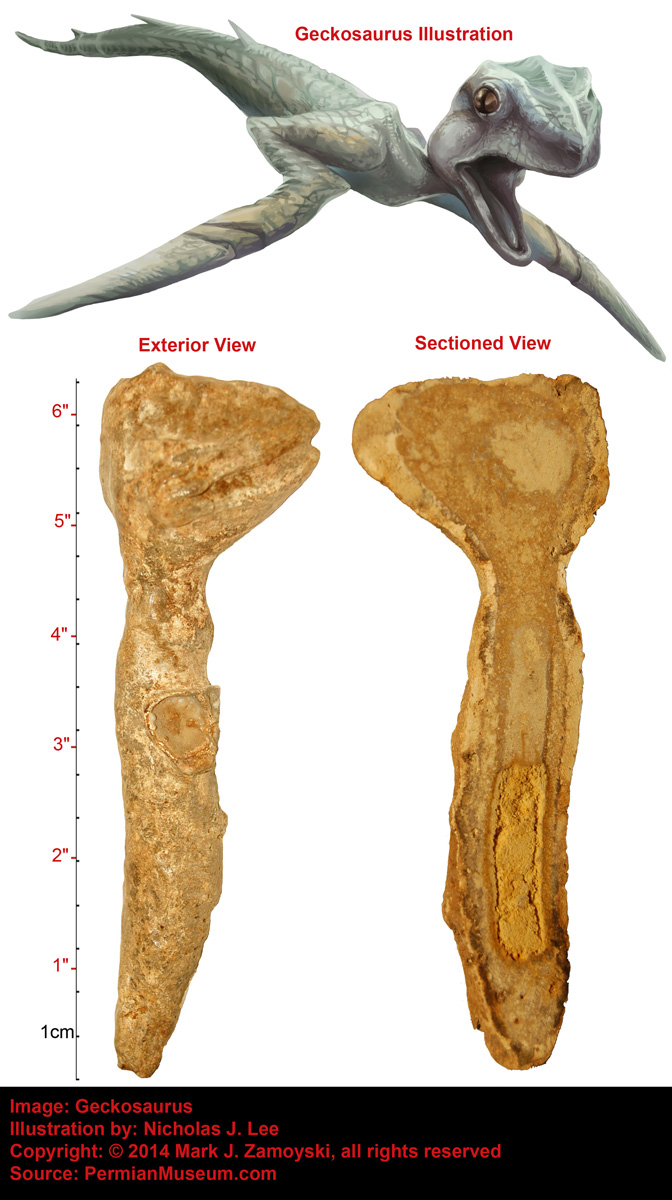
Lizard DNA is extremely important in the origins of Dinosaurs. Dinosaurs first appeared around 250 Ma, or right after the last quantum re-speciation / recovery pulse of the Permian period. The word Dinosaur itself derives from the Greek deinos "terrible" plus sauros "lizard".
Birds
Bird like life forms, preserved as whole body limestone fossils, are fairly common in our collection.
However, our initial focus will be on combinational life forms because of their significance in corroborating the genetic reassortment process and potentially serving as the aquatic forerunners of birds.
Birds may have originated from aquatic life forms that used "water wings" for propulsion. Several of the fossils appear to be aquatic life forms that are morphologically similar to birds, with the major exception that they did not have feathers but skin more suited for the ocean.
One such example is Aqua Duck, which is morphologically similar to a duck, except without the feathers and extensive wing structure necessary for propulsion through air. However, the propulsion structure would be more than adequate for propulsion through water.

Genetic reassortment could also be expected to generate hybrid fish / bird life forms, in a manner similar to the fish / lizard combinations discussed in the previous section.
An example of such a fish / bird "Amalgamation / Combinational Life Form" (or fish / insect combination) is the aquatic life form called Birdsquito, which has the head of a bird and the body of a fish. A close up of the beak from both sides is shown below and reveals one side has serrations in the beak (Side B - Headshot Zoom) or possibly a filter feeding beak.

Another example is an aquatic life form that looks like a mix between a Seahorse and a Bird. The vestiges of a "water wing" run vertically down the center of the life form. The head and beak are similar to those of aquatic birds. This may be an early aquatic forerunner of birds, and birds in turn appear to also be the forerunners of some terrestrial uprights.

The specimens above were presented because of their significance in corroborating the genetic reassortment process as well as providing an insight into possible aquatic forerunners of birds.
However, just like with the fish, there are also a larger amount of "normal' bird fossil specimens.
Removing the matrix from the upper part of a longer necked coastal bird specimen shows the crystallization is so good that it reveals external features such as the eye and beak of the life form. The bird appears to be more akin to a "normal" bird, rather than combinational life form.
External View of Long Neck Bird Head and Neck
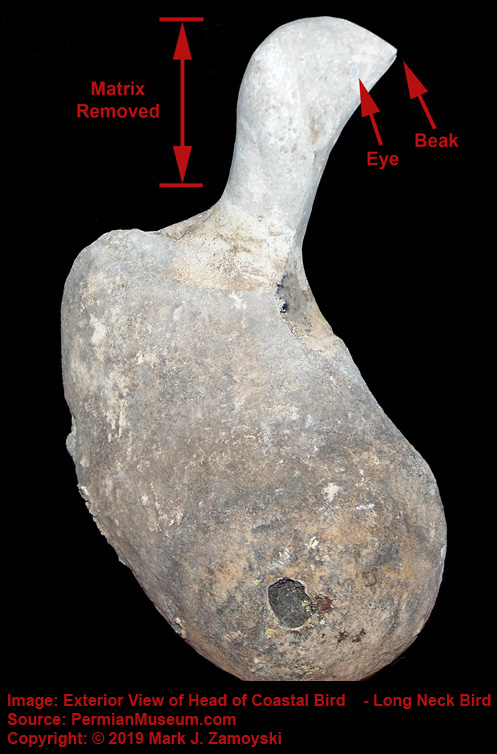
Sectioning another specimen also reveals a more normal "bird only" life form we call Short Beak Bird. It should be noted that this specimen appears to have the integrated CSFF DNA in both the head area and wing area, similar to the Pteranodon Pelican specimen, which is presented and discussed at the end of this section.
Sectioned Small Coastal Bird
Birds are often found near fossilized eggs. Cutting one of the eggs open reveals an internal developing infant bird. It should be noted that the egg has a cementing layer attaching it to the limestone substrate.
Sectioned Fossilized Egg Cemented to Substrate
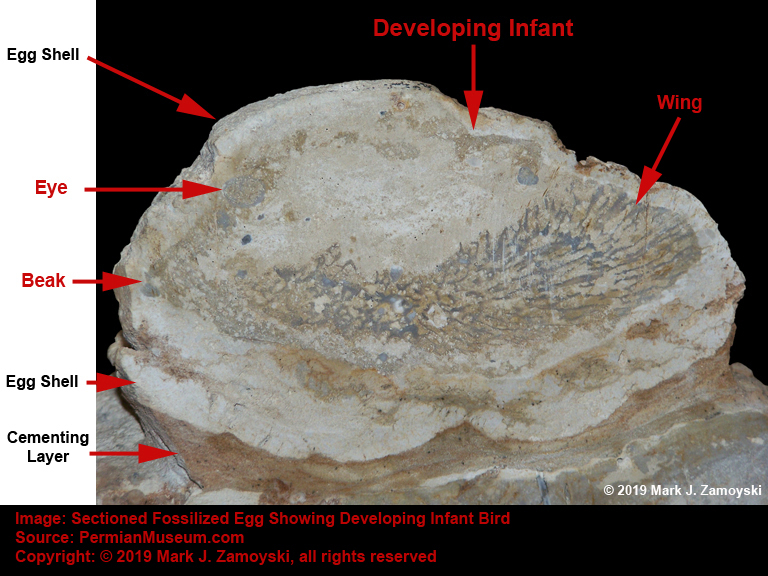
The cementing may have been done to keep the egg from being washed away. Flamingos today still use elevated mud nests to protect their young for rising and falling water levels.
However, it may also have been done to protect the egg from being eaten by the mud worms that lived below. Larger eggs in particular appear to have extensive worm burrow activity below. We have recently found a fairly intact specimen of one such egg eater worm, Mud Worm 2, which is shown below.
Mud Worm 2: "Permian Graboid" Below the Eggs

The subterranean worms that lived in those burrows are covered later in the Worms and Insect Section of this chapter.
It appears some birds also ate infant and juvenile worms. The sectioned specimen we call WormEater Bird appears to have linear worm-like life forms and segments in its stomach. Alternatively, this may also be a bird with a parasitic nematode infestation.
Worm-Eating Bird

Birds eventually went on to master flight and moved their eggs to nests in trees and nests on hard stone cliffs, where the "Permian Graboid" type worms could not reach them.
Birds like today’s Robin still seek out and eat earthworms.
There are also birds in the collection that appear to display the head structure of what would become the Pterosaurs (wing lizard), but without air wings yet. An example is shown below appears to have a head like the early pterosaurs, but no wings and a possible contractile sack propulsion system (bottom of sectioned specimen):
Likely Wingless Aquatic Forerunner of Pterosaurs
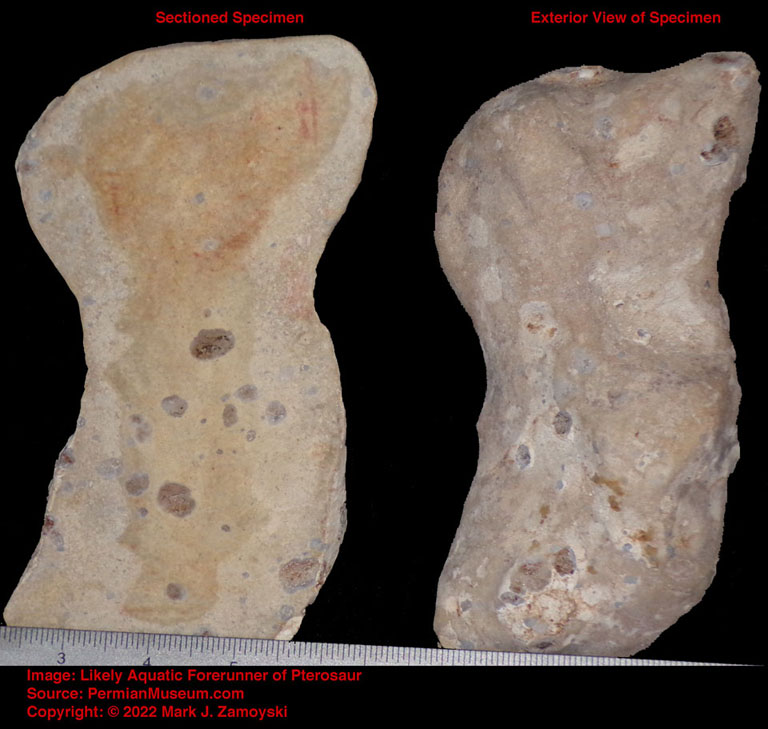
"Air wings" would require skeletal or boned structures, which leads us to a likely successor to the specimen above, covered in the Pterosaur / Pteranodon lineage in the Pre-Dinosaur section.
Pre-Dinosaur DNA
Dinosaurs are a diverse group of animals that first appear in the fossil record right after the last Permian recovery (i.e. quantum speciation) pulse. Their features are consistent with the DNA available for that last Permian recovery pulse, as will be covered in this section. The specimens in this section fall into the DNA reassortment prediction of " Future Life Form Forerunners".
Large marine animals called Pistosauroidea first appeared between 252 to 247 Ma.
Large terrestrial animals called Pseudosuchia (such as the Poposaur) appeared 250 Ma.
Large birds capable of flight (such as the Pterosaurs), appeared around 228 Ma.
Starting with the large birds:
"Air wings" did not appear until around 228 Ma with the Pterosaurs. Pterosaur wings were not feathered but formed by a membrane of skin, muscle, and other tissues that stretched from a simple rod like bone structure at the leading edge and were attached at the rear to the ankles, as shown below:

The specimen we call Pteranodon Pelican displays what appears to be two integrations of CSFF DNA, one in the head that likely went on to become a beak and skull, and a skeletal matrix in the wing area that likely went on to become the rigid boned wing structure required for airborne flight.
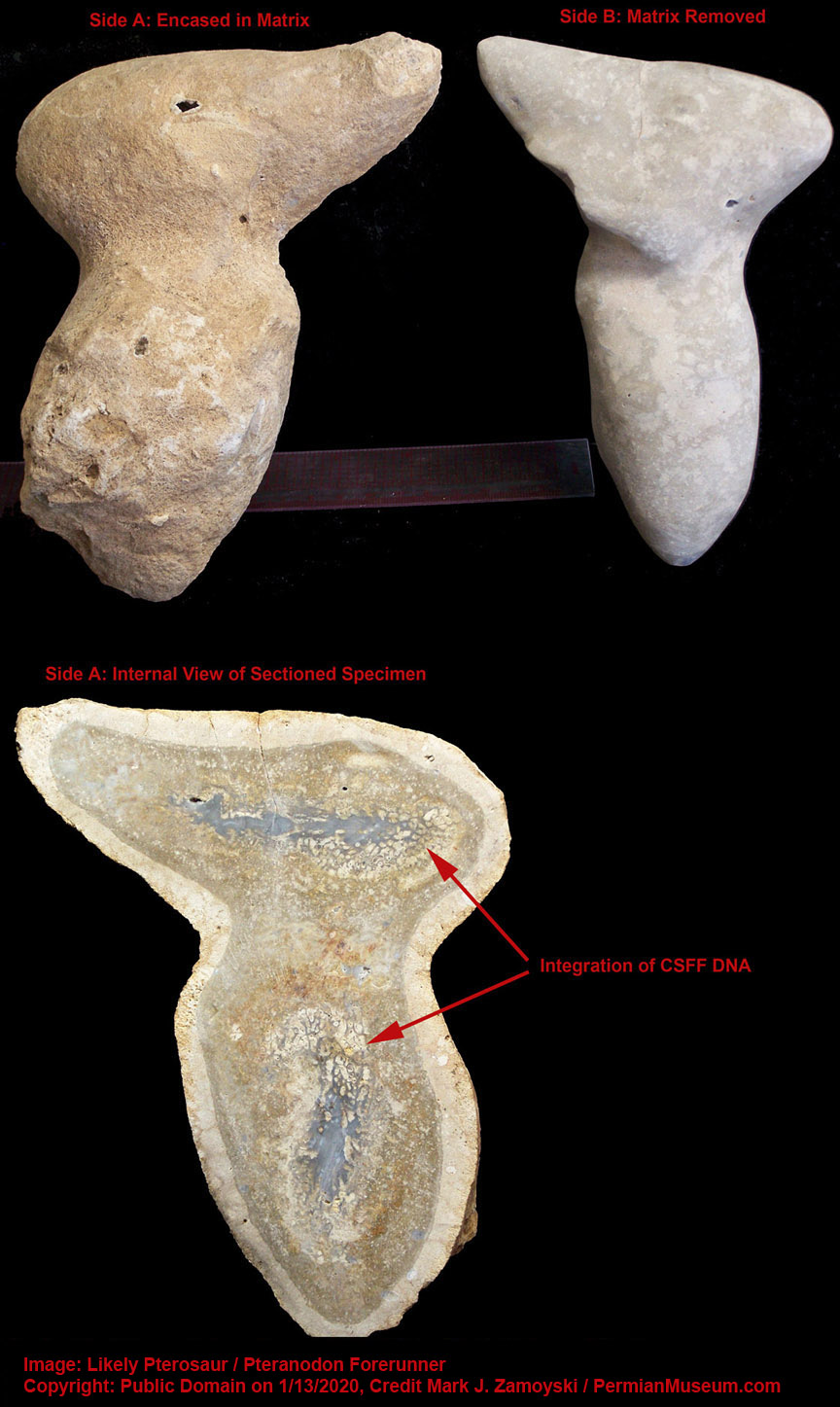
The CSFF structures appear to follow the linear growth pattern observed in many of the stand alone CSFF structures previously presented. In the absence of growth control pathways, the projected growth of the newly integrated CSFF skeletal structures would be as, shown below:
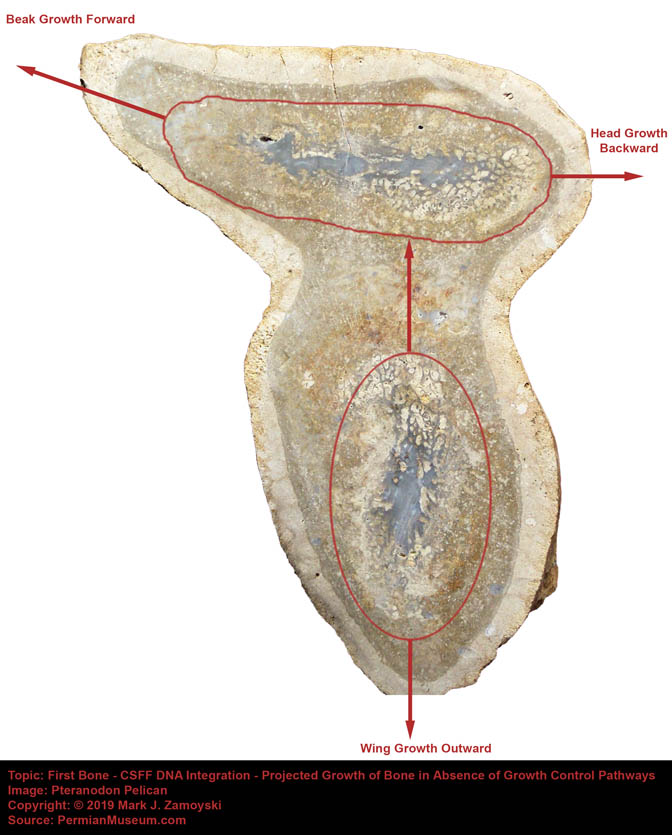
The Pteranodon’s exaggerated beak growth forward and head growth backward, and extended wing bone growth outward, are consistent with the projected linear growth pattern of the integrated CSFF DNA in the Pteranodon Pelican specimen.
The importance of bone in the advancement of life goes well beyond just its role in skeletal function or allowing for air flight. Bone is a repository for Calcium, Phosphorous, and Mitogens (growth factors). These compounds are routinely moved from extracellular fluid into bone and back from bone into extracellular fluid. Calcium (Ca++) movement alters nerve function, muscle function, consciousness, and memory. Movement of mitogens, Ca++, and phosphorous activate the population density management / cell cycle control system. This is an extremely significant advancement in life and will be discussed extensively in a later chapter.
Raptor DNA appears to have gone into many lineages, including the Eudromaeosaurians (e.g. Velociraptors, Deinonychus, Utahraptor, Bambiraptor) that inhabited earth from 143 Ma - 66 Ma. These life forms looked like they were part bird and part reptile.
The combinational bird/reptile DNA lineages (e.g. Velociraptors) perished during the K-T extinction, likely as no terrestrial animals over 25 kilos (55 pounds) survived the global cooling that followed.
The smaller bird (raptor) DNA has survived to this day. As a contemporary example, Arizona's Harris's hawks are pack hunters comprised of a dominant female and two males. They basically prey on anything smaller, including birds, mammals, and reptiles.
Harris's Hawk Breakfast: Dove, With a Side of Starling

As for the large marine animals:
One of the earliest large marine animals appears to be a winged dragon-like life form that has an over sized brain cavity.
Winged Nematode with Brain Cavity
Specimens in the collection (e.g. DinoSeal) have multiple neurons (optic, olfactory, and vibration sensing) that connect to different muscle groups. Routing them through a brain would now allow coordinated responses to environmental stimuli.
More importantly, a brain and brain cavity allowed for integration and hyper-proliferation of glial cells. Astrocytes are the most numerous glial cell. They interconnect neurons and are responsible for propagation of the calcium ion wave in the brain which hosts the measurable electromagnetic phenomenon underlying consciousness and memory formation.
As an example, neurons make up 97% of a Leech brain. In contrast, glial / astrocyte cells make up 90% of a human brain, and only 10% are neurons.
The appearance of a brain cavity that was much larger than necessary to house just neurons suggests glial cell content hyper-proliferation, which was a significant advancement toward life becoming "aware".
One of the first dinosaurs to appear was a marine animal called Pistosauroidea, which first appeared around 252 Ma. Pistosauroidea went on to spawn a long lineage, including the Plesiosaur, which first appeared around 203 Ma. Plesiosaur became common during the Jurassic Period ( 201 Ma to 145 Ma) and eventually went extinct around 66 Ma during the Cretaceous - Tertiary extinction event. Plesiosaurs were air breathers that bore live young.
An illustration of a Pistosauroidea is provided for reference below, as our fossil collection includes forerunners of the Pistosauroidea, and hence forerunners of the Plesiosaurs.
Pistosauroidea - Forerunner of Plesiosaurs

Our fossil collection includes 2 specimens that appear to be forerunners of the Pistosauroidea / Plesiosauria dinosaurs.
The first is a sectioned specimen of what appear to be two infants, preserved as the bluish translucent crystals previously discussed. The infant on the right provides a better view of what the life form looked like.
Pistosauroidea / Plesiosauria Forerunners (specimen 1 of 2)

The second specimen is the external view (with matrix removed) of a juvenile. The juvenile appears to have died as the result of being chomped by something, as the right front side of the life form (left side of the picture) is flattened and a somewhat circular jaw impression is visible starting at just below the head and extending halfway down the body. The specimen appear to only have two flippers, not four.
Pistosauroidea / Plesiosauria Forerunners (specimen 2 of 2)

As for the forerunners of the forerunners of the Pistosaurs, there are two specimens in our collection that appear as potential candidates. Just as DNA reassortment can result in life forms like Double Fish previously reviewed, the integration of two flippered life forms, could result in a four flippered life form such as the Pistosaur.
The front end of Pistosaur would be consistent with a specimen we call Dino-Seal.
The nose would suggest Dino-Seal was an air breather like the Pistosaurs.
Dino-Seal appears to have an olfactory (chemotaxis) neuron, in addition to an optic (phototactic) neuron, as well an unidentified neuron. All three appear to be connected to a different motility group. There is no evidence of a brain.
In the absence of a brain, it is unclear how the life form would react in situations where there was a conflict among the three neurons. However, the presence of a nose and nasal passages, with the olfactory bulb positioned at a gap in the nasal passages, strongly suggest Dino-Seal was an air breather and the olfactory neuron would likely only be active when the animal was above the surface of the water and breathing. This would preclude any potential for underwater conflicts with the other two neurons.
In context of an air breather, the optic neuron would likely function to drive the air breather to the surface, as moving toward light would mean moving toward air.
If the unidentified neuron is an auditory sensor, it would more likely have been specific to vibrations in the ocean, as there is no orifice for air to enter (i.e. no ear canal). In an underwater context, move toward vibrations would mean move toward food.
Accordingly, the only conflict underwater would be 1) move toward food, or 2) move toward air. It is not clear how this would be resolved without a brain. It may have simply been on which of the two signals was stronger.
Dino-Seal only had two flippers. If a DNA reassortment resulted in a duplication (akin to double fish), where a second set of fin DNA was appended to the back of Dino-Seal, this could yield a four flippered life form variant of Dino-Seal. The DNA for the rear flippers would not even have to be related to Dino-Seal, and actually it appears to be morphologically more similar to that of the Stingray. A photograph of the Stingray rear is juxtaposed next to the Pistosaur Forerunner photo to illustrate the point:
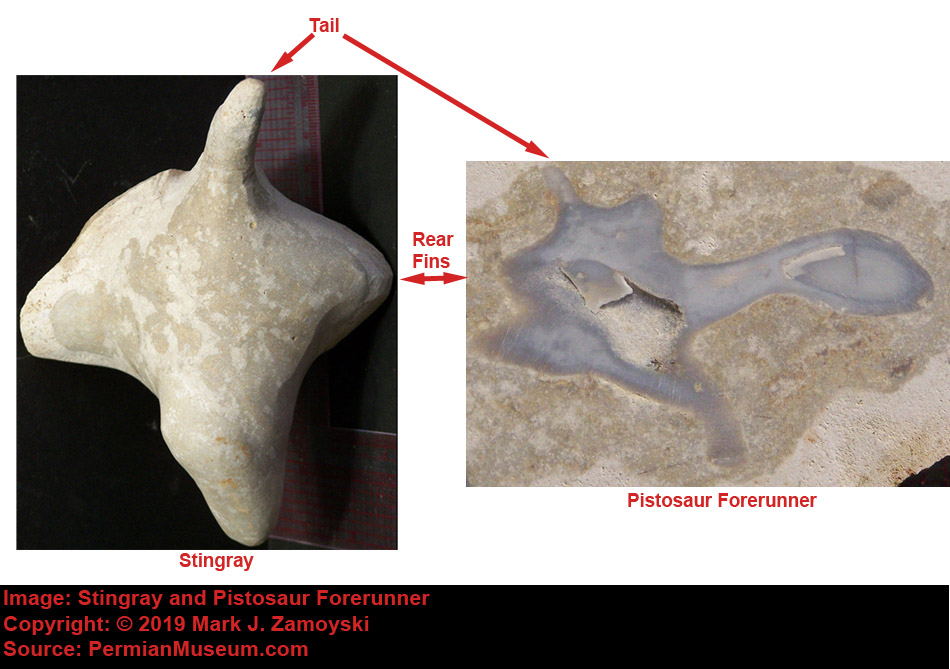
A DNA reassortment that creates a life form that has Dino-Seal as its front and a Stingray’s rear is just one of innumerable possible reassortments that could result in a Pistosaur Forerunner with four flippers instead of two.
As for the large terrestrial tetrapods:
Terrestrial tatrapods may have come in part from the air breathing, morphologically similar aquatic counterparts, the Pistosaurs.
There were two specimens found with the Pistosauroidea that appear to be early terrestrial tetrapod forerunners, possibly DNA reassembled versions of the Pistosauroidea, however also possibly of unrelated origin.
The first is a sectioned specimen of an infant, preserved in the bluish translucent crystal type of fossilization. We have named it Tetrapod Plesi, as it was part of the same stone from which the Pisto / Plesi forerunners were sectioned.
Tetrapod Infant Found with Pistosauroidea
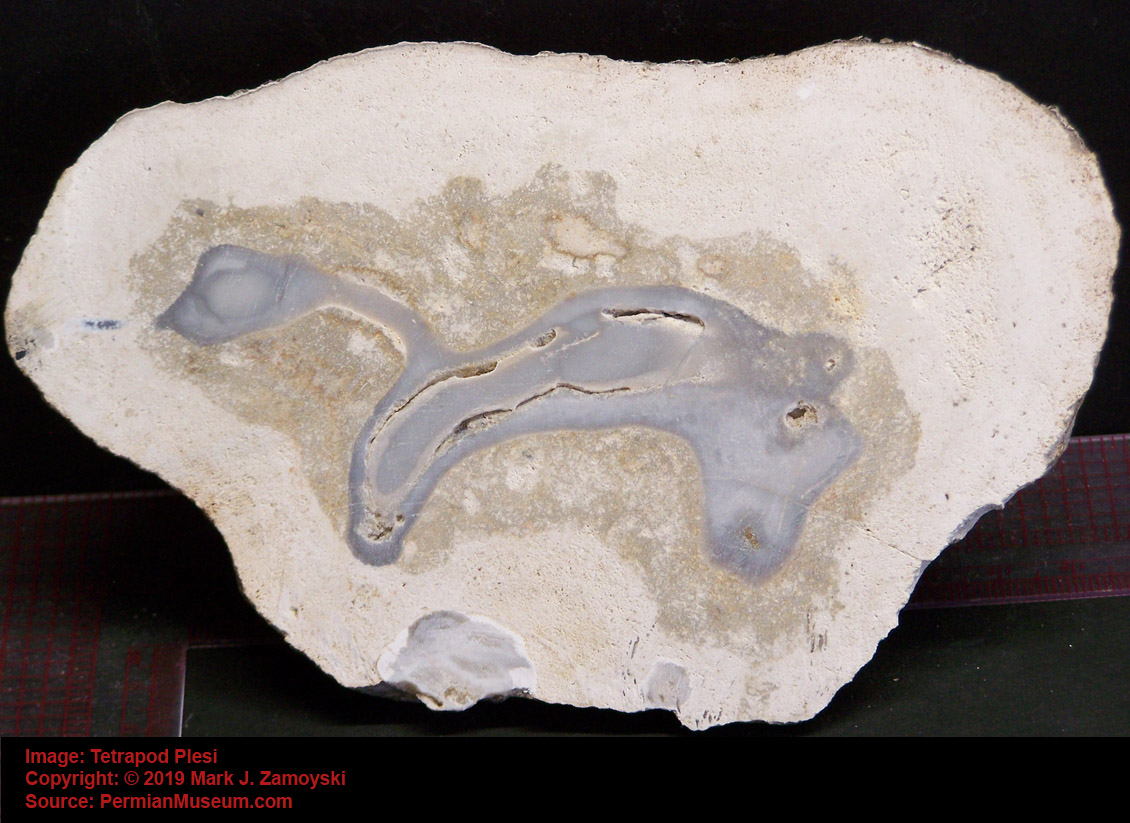
The second Tetrapod Plesi specimen appears to be a juvenile and is still in matrix pending processing (shown below, in matrix):
Tetrapod Juvenile in Matrix (Pending Processing)

Brontosaurus, or "thunder lizard", did not appear on land until around 156 Ma, however it appears its DNA may have come in part from the Pistosauridea (252 Ma) or its decendent Plesiosauria (~ 204 Ma to 66 Ma).
Brontosaurus Illustration

A photo of the Bronto-Pistosaur fossil is shown below:
The photo shows a sectioned view of the specimen that is ~ 6 inches high and 3 inches wide. On the central, right side of the photo, there was a slight air gap between the fossil and the matrix. This allowed us to remove this part of the matrix from the fossil to reveal the body style more completely.
Bronto - Pistosaur Fossil Photo

As for terrestrial uprights that walked on two feet:
It appears that the 250 Ma Poposaur was an upright that acquired most of its DNA directly from the lizard lineage and required no unusual forerunners. Reptiles were among the survivors of the end Permian extinction and were the likely antecedent lineage for the large land lizards. Because our fossils come from a shallow coastal ocean, no large terrestrial animals could be expected in our collection.

Poposaurus was basically a large land lizard with upregulated growth (or absence of growth control pathways). It displays diminutive front limbs, indicating it may instead be a forerunner of some of the upright walking lizard like terrestrial life forms that had two legs and two arms (e.g. T-Rex).
Reptile on a Rock
DNA reassortment of DNA from lizards, birds, or like forms like Geckosaurus would provide innumerable upright terrestrial life form possibilities.
While the Poposaurus appears to be a transition from four legged to two legged lizards, birds appear to have been forerunners of velociraptors and other upright life forms.
Based on our "coastal" fossils, the birds spawned during the Permian explosion of life appear the first of the new life forms that emerged from the ocean to walk upright on land.
An example of a walking bird is shown below, and its legs are shorter and more stocky (red arrows) than most birds today.
Walking Bird

The collection also includes uprights that appear to have structures for hopping rather than walking.
The first of these upright "combinational" life forms appears to have the head and beak of a bird but a body more closely resembling a kangaroo. The much smaller tail appears to be a transitional feature often observed in the move to upright mobility.
Indeterminate 3: Upright with Kangaroo like Body
The next combinational life form can have a dozen possible interpretations, so author will limit its description to "one of the early uprights", with a diminutive tail characteristic of the move to upright mobility.
Indeterminate 41: Early Upright
Important Caveat: We will continue to add uprights as we find them, however keep in mind our fossils are limited to a shallow coastal ocean and nearby coastal land during the Permian period. The fossils are not from deep inside land masses. That pretty much limits our fossils to 1) early forerunners of what eventually went inland and 2) is limited to life forms that were spawned during the Permian Explosion of Life.
Worms and Insects
The fossilized specimens of worms and insects in the collection are typically 6 inches or more in length. However, in the absence of established growth control pathways, the size of adults would only be limited by availability of nutrients.
Worms
Starting with the worms, we previously mentioned how there are numerous eggs in the collection:
And under some of the larger eggs is fossilized evidence of extensive worm burrow activity:

We have finally found a fairly intact specimen of the subterranean worm that lived in those burrows and presumably ate the eggs. We have named the specimen the "Permian Graboid", the term Graboid being coined in the movie "Tremors" which featured a similar prehistoric life form.
Subterranean Egg Eater Worm
The Permian Graboid appears to have a set of lower and upper mandibles.
The lower mandibles appear to be horizontally oriented pincers similar to those one may see on a bull ant, stag beetle, or cheiracanthium punctorium spider. Horizontal insect pincers are typically used to grasp, crush, or cut food.
The upper mandibles of the subterranean worm are vertically oriented with a right angle bend, possibly jointed, and similar to a claw hammer or angled fangs. Spiders have fangs, first appeared 386 Ma, with upgraded versions appearing 318 - 299 Ma, so the DNA for both the lower and upper mandibles was available at the time of reassortment. Some species of mantis shrimp, which first appeared 340 Ma, have similar jointed structures they swing upwards to break shells. The claw hammer of the worm would likely have been pulled down, instead of swung up. An illustration of what the mandibles look like based on the newly discovered Egg Eater Worm:
One possibility is that the Permian Graboid’s lower pincers were used to push through the shell and hold on to the egg, while the upper claw hammers or jointed fangs were used to punch a hole in the egg shell and then pull the contents of the egg into the Graboid’s mouth.
One of the original 35 fossils was a worm that had a chitin like exoskeleton, but was missing the mouth. The illustrator took some artistic license to create a mouth, which we now know is uncannily close to reality. It appears the first "Mud Worm" specimen, or Mud Worm 1, was effectively an armored graboid.

A zoom of the tail show a better view of the exoskeleton.

Worms today are generally soft bodied and insects are characterized by a chitinous exoskeleton. In a DNA reassortment world, traits are readily shared between unrelated life forms via the DNA reassortment process, and categorizing life forms by today's standards is not always possible.
The most significant worm discovered is "Brainiac Worm", which has 3 separate integrations of CSFF DNA, one in the head forming a skull and two in the tail functioning as a "jointed" limb. The life form was effectively a head, pushed along by an attached jointed finger.
An illustration and sectioned specimen photo of Brainiac Worm, with a zoom of the tail is shown below:
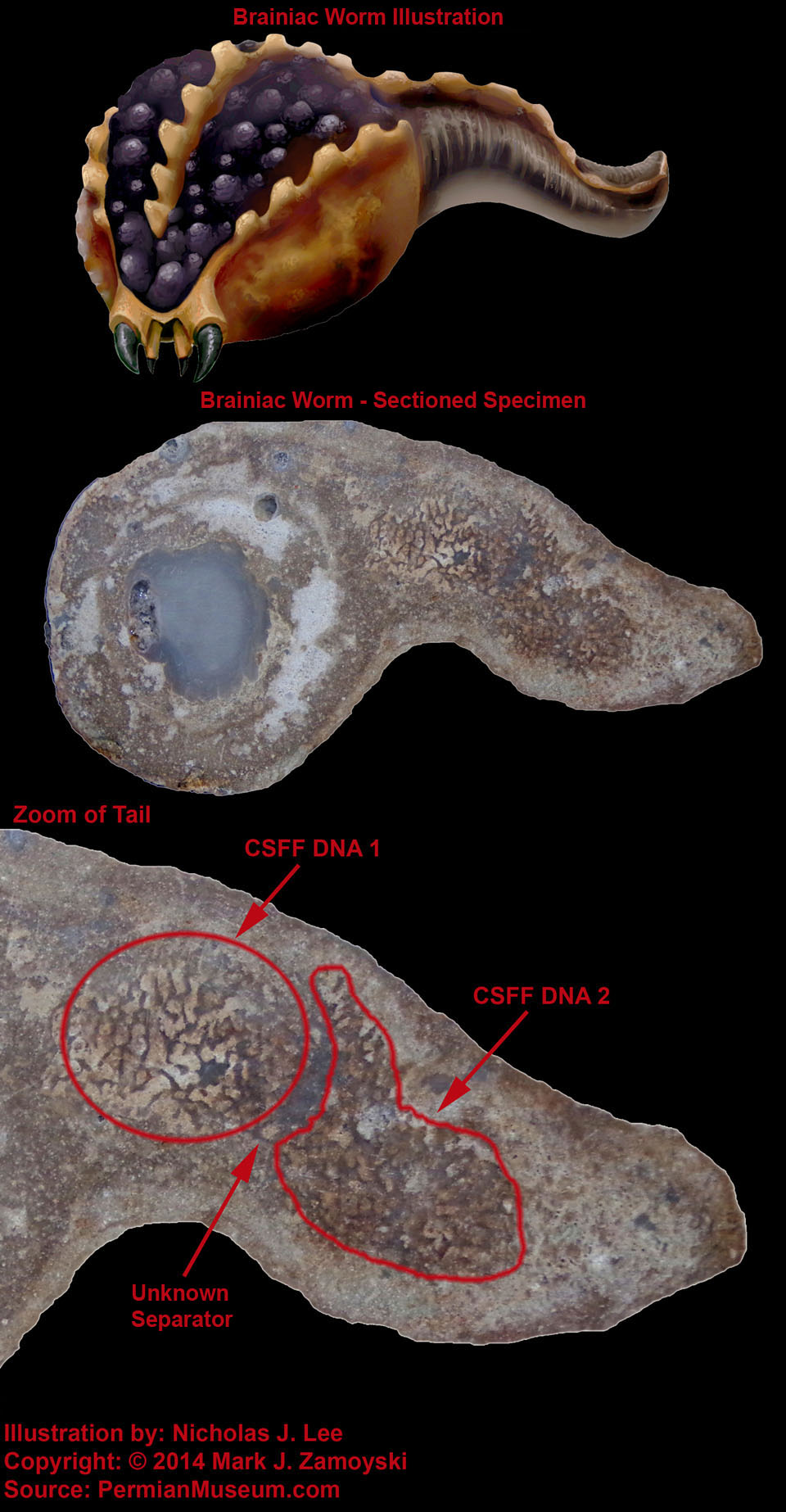
The zoom of the sectioned tail shows an "unknown separator" between the two CSFF DNA structures, allowing for jointed motion, or "mobility" versus just "motility". The arch of the tail would not be possible without the ability of the two hard masses to slide past each other, which is what the unknown separator appears to allow.
What generated the separator (e.g. the soft tissue life form, or the CSFF structure(s), or something else) is not known at this time. What is know is that this is the first reassortment specimen in which we have observed "jointed bones" .
Insects
The insects that are part of the fossil collection are not like insects as we know them today. Much like the fish / lizard combinations the winged insects are more akin to insect / bird combinations. They are also much larger than today, typically 6 inches (25 cm) or longer in length. Some have a venom sack in the rear with the injection needle pointed upwards and to the back, indicative of a more defensive mechanism against being eaten, rather than the more offensive / defensive bottom mounted, forward facing stingers in use today.
The first specimen is called Hornet Hawk and appears to have a head closer in appearance to that of a hawk, with a body more closely resembling a hornet, with the notable exception the stinger is pointed upward, rather than downward.
Insect / Bird Combinations - Hornet Hawk
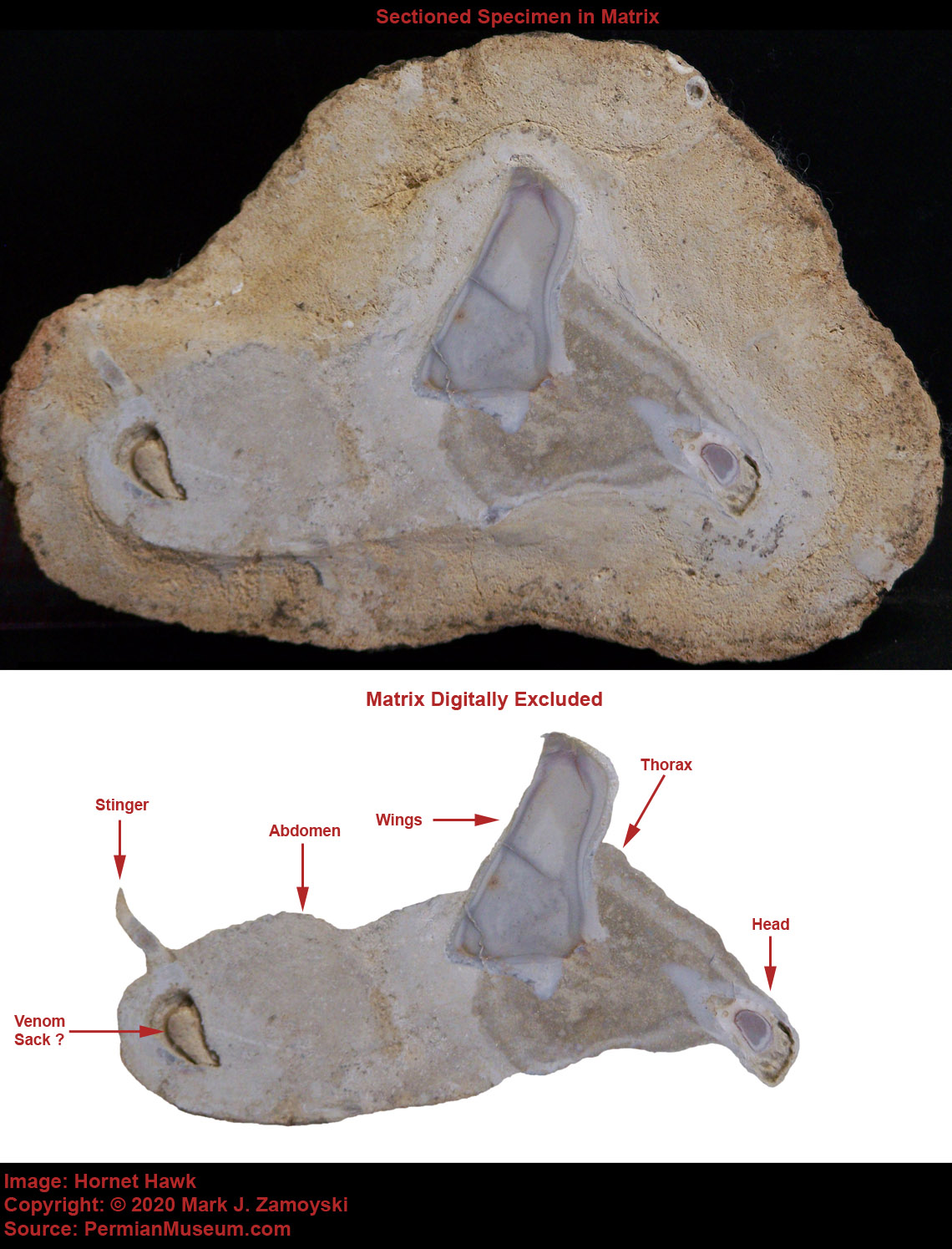
The second specimen appears to be a bee / bird combination because of the CSFF integration in the wing. Bees have membrane wings without bones:

CSFF integration with soft tissue has been observed as a "bone forerunner" in bird wings, such as the Pterosaurs previously examined.
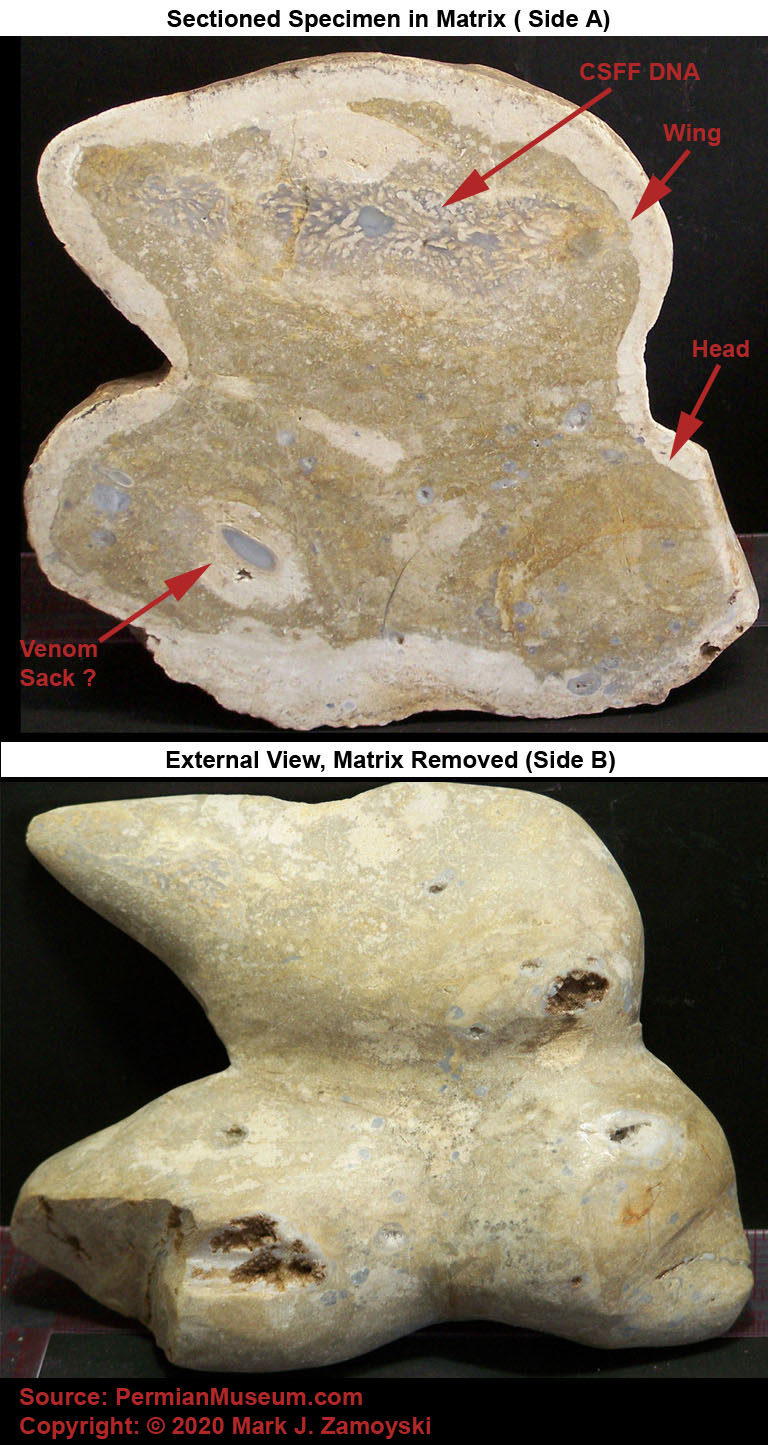
The bee forerunner appears it may have a venom sack and upward stinger (see sectioned specimen photo), similar to that of Hornet Hawk. The mouth of the specimen ( see external view photo) appears to be neither bird nor insect, but more akin to lizard or fish.
The upward facing defensive stinger at the rear is not unique to the insect specimen. It also appears in fish specimens, as previously covered.
CSFF Hybrids
Life on earth starts with a single cell. All that cell needs is a nutrient rich environment (egg, womb, petri dish, etc...) and it will relentlessly grow and divide to become the life form coded for by its DNA. At a point in its maturity, it will be able to leave that nutrient rich environment and actively pursue food in the surrounding environment.
It appears the CSFFs may also have provided a nutrient rich environment, in and around the CSFF, for newly created cells to thrive in, somewhat akin to a petri dish. It also appears some of the newly created cells never left.
While the fossils presented so far are single organisms that posses the DNA and features of multiple organisms, the fossil specimens in this section are "Biotic Hybrids", and more specifically CSFF / Soft Tissue biotic Hybrids. A hybrid is a thing made by combining two elements. In this case, the underlying organism is a CSFF and the other organism resides on or in the CSFF.
Since CSFFs shred suspended cells, they are repositories of readily available nutrients. It is not surprising that soft bodied organisms would establish residence on or in CSFFs, either for structural attachment purposes or because of the proximity to nutrients. These relations can be symbiotic or parasitic.
The first example is a CSFF / Soft Tissue hybrid that appears to be a small carnivorous CSFF. The top part of the CSFF has a pincer like mouth that may have the ability to actively capture multicellular life forms, rather than just filtering feeding of single celled organisms.
CSFF Hybrid 1: Pincered CSFF
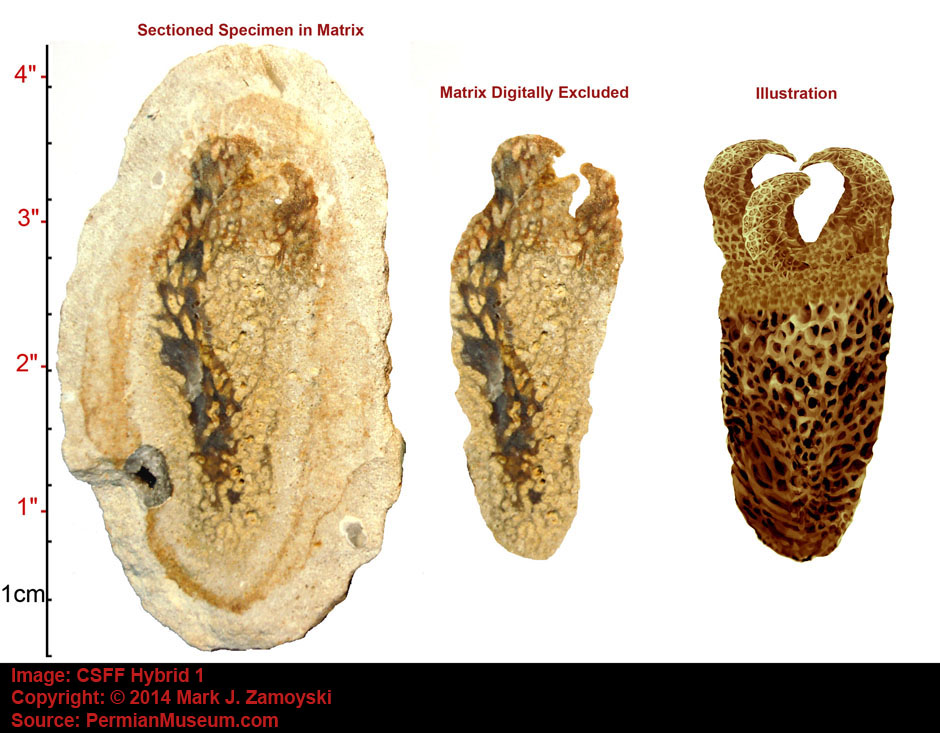
The second example is a larger, more pincered CSFF that starts looking more like a Venus flytrap. A Venus flytrap has hinged leaves that spring shut to capture insets that land on them.
CSFF Hybrid 2: More Advanced Pincered CSFF
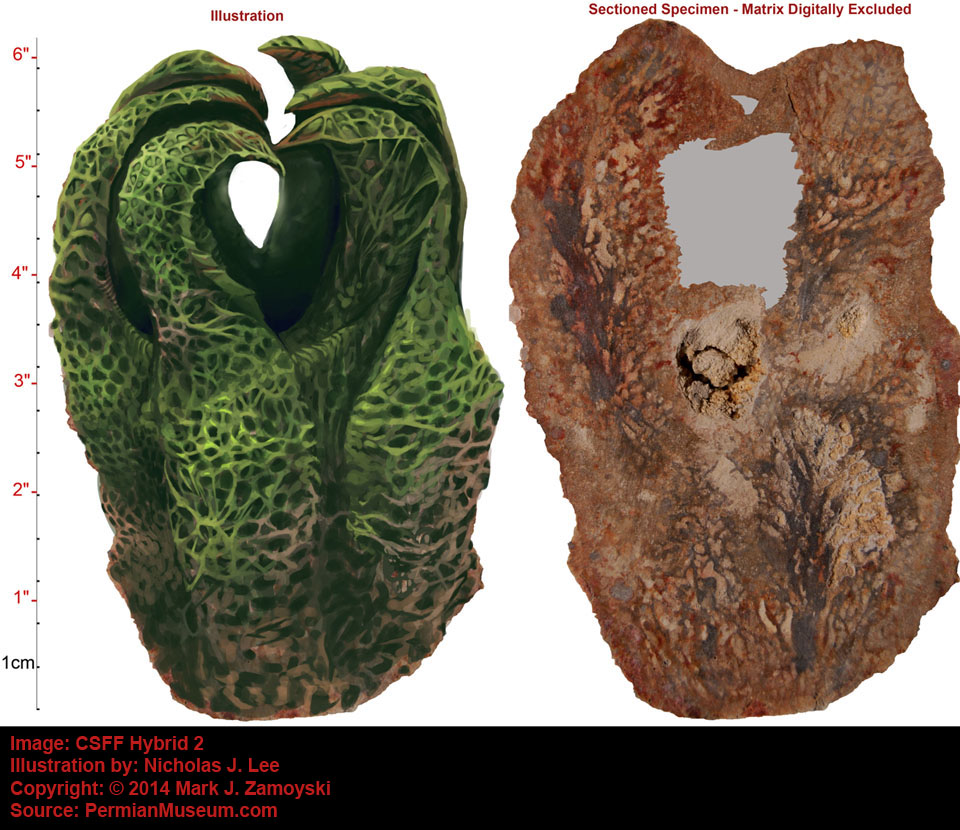
While the first two CSFF Biotic Hybrids may have been symbiotic, the next ones seem to be more parasitic and detrimental to the survival of the CSFF colony. CSFF Hybrid 3 appears to be a CSFF encased by a surrounding coat of cells, that would prevent water from entering the CSFF and allowing it to filter feed.
CSFF Hybrid 3: Encased CSFF
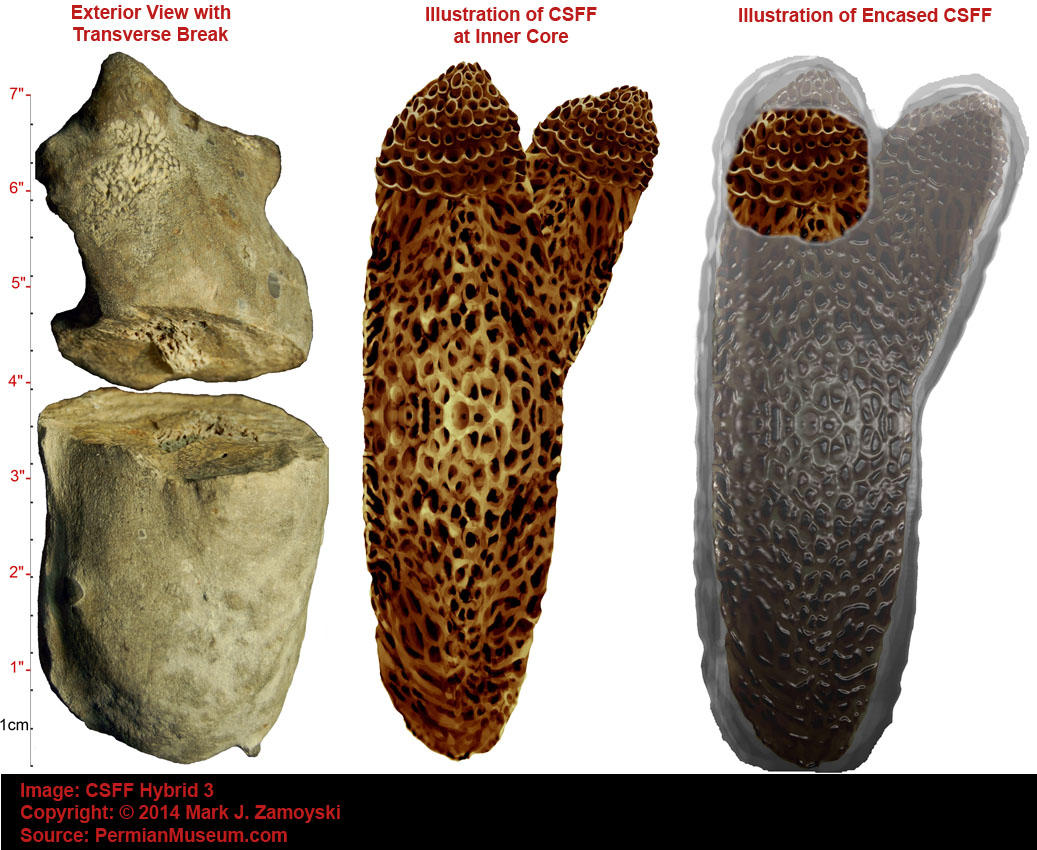
While all of the above CSFF Hybrids have the soft tissue on the outside, CSFF Hybrid 4 has a soft tissue life form on the inside. While the soft tissue life form benefits from the nutrients on the inside, it also prevent the CSFF from being able to filter feed. The illustration shows the eventual predicted demise of the CSFF, as the soft tissue life form within the CSFF continues to grow.
CSFF Hybrid 4: Internal Parasite

Chapter 5
Life Science Implications of the Specimens
Copyright © 2012 Mark J. Zamoyski. All rights reserved.

Physiological Life
A feature common to all motile multi cellular specimens is that they have a “Skin” that surrounds the collection of eukaryotic cells and allows the cells to be bathed in fluid. Colonies of single celled organisms have the ability of form a protective outer layer of cells around the colony. Skin may have originated from this DNA.
In context of multi cellular eukaryotic life, skin is the most significant development for several reasons:
Evolution onto Land: The ability to encapsulate and control a microenvironment set the stage for migration from the oceans onto land. A 70 kg human has ~ 40 liters of extracellular fluid bathing the cells. Humans effectively needed to take their gulp of the ocean with them before they could step onto land.
Protection of Eukaryotic Cells: Eukaryotic cell walls are more fragile than prokaryotic cell walls and the encasing fluid distributes various external traumas over large areas, preventing rupture of the eukaryotic cells walls.
Cell Signaling: A skin encased aqueous microenvironment took cell signaling to a whole new level. Autocrine, paracrine, and endocrine molecules are not washed away by the ocean but are now contained in a microenvironment where they can much more effectively reach their intended target cells.
Ion and Concentration Gradient Maintenance: In this new microenvironment, concentration gradients could be controlled to support multi cellular life. Physiological life as we know it is basically a collection of concentration gradients. Presence of concentration gradients means life. Absence of concentration gradients means death.
The aqueous microenvironment allows atoms to be maintained as free ions. As an example, sodium (Na) loses the single electron in its outermost valence shell and acquires a positive charge, becoming a positive ion designated as Na+. Calcium loses two electrons becoming Ca++ . Chlorine (Cl) has 7 electrons in the valence shell and gains an electron becoming a negative ion designated as Cl-. Without water, sodium and chlorine combine into the neutral molecule NaCl (table salt).
Ions are electrically conductive. Concentration gradients of ions are called electrochemical gradients. Traveling perturbations in electrochemical gradients carry signals along the axon (long body) of a nerve cell to the synapse, where they are then converted into a chemical signal.
Nerve Function: Electrochemical gradients allow for function of a cell called a neuron. The most rudimentary function of nerves is perception of the environment and initiation of a coordinated response to the environmental stimuli. In more advanced organisms, neurons play a part in consciousness and memory formation.
A neuron is a single cell. The specialized task of a neuron is to receive, conduct, and transmit signals. Sensory neurons, motor neurons, and interneurons all have the same overall structure: a spherical central cell body (soma) that contains the typical organelles found in all cells, branching dendrites on one side to receive signals, and a long axon on the other side for transmitting signals. The axon commonly divides into many branches at its far end so it may pass the message to many target cells simultaneously.
Neurons contain ion channels that maintain an electrochemical concentration gradient balance between ions (primarily potassium, sodium, and chloride) so that the resting membrane potential inside of the neuron is around -85 mV relative the outside of the cell (ranges from -30 mV to -100 mV depending on cell type). Changes to the membrane potential are called “depolarizing” if they make the inside of the cell less negative. Depolarizing a nerve makes it much easier for the nerve to fire (i.e. lowers the input voltage required for the nerve to fire).
Above a threshold level, a depolarizing event initiates a traveling perturbation in the electrochemical gradient, which travels until it reaches the end of the axon which terminates in structures called synapses.
Propagation of initiated perturbation toward synapses at the end of the axon is shown below:
Each synapse contains transmembrane voltage gated calcium channels and intracellular vesicles containing neurotransmitter.
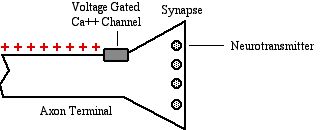
When the electrochemical signal reaches the synapse, the voltage gated calcium channels open transiently, allowing an inrush of calcium ions (Ca++).
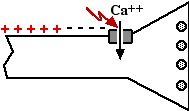
In inrush of calcium ions causes the vesicles containing the neurotransmitter to fuse with the synaptic cell membrane and release the neurotransmitter into the post synaptic gap (aka cleft).
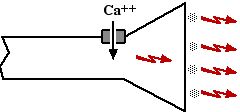
The intended target of the neurotransmitter depends on the type of the nerve and what it is connected to.
As an example, if the neuron is connected to muscle (neuromuscular junction), the release of the neurotransmitter acetylcholine causes the muscle to depolarize via neurotransmitter gated channels. The depolarization spreads along the muscle surface and the T-tubules that run along the surface of the muscle fibers. The depolarization opens voltage gated Ca++ channels in the T-tubule surface that allows Ca++ from the extracellular fluid in the T-tubule to enter the the sarcoplasmic reticulum. The inrush of Ca++ into the sarcoplasmic reticulum activates the “sarcoplasmic reticulum calcium release channels” (SRCaRCs), which in turn release Ca++ into the fluid around the myofibrils. The released Ca++ allows the muscle to contract by removing the tropomyosin block between actin and myosin, triggering cross-bridge formation by enabling myosin to bind to actin.
The Bone Micro Environment:
Normal bone undergoes a continual remodeling process that essentially replaces the entire skeleton every 10 years. Remodeling is mediated by two cell types, osteoclasts which dissolve bone (resorption), and osteoblasts which are the bone builders. Both cell types come together in three to four million sites scattered throughout the skeleton. During childhood and adolescence, bone formation proceeds at a faster rate than resorption. By around age 40 bone resorption begins to outpace bone formation and bone thinning begins to manifest.
Cyclicality of bone destruction followed by bone building appears to be an important aspect required for maintenance of bone density. Intermittent administration of parathyroid hormone (PTH), which increases osteoclast activity and bone destruction, results in an eventual increase in bone mass, whereas continuous administration of PTH results in eventual bone loss. The point being, while continuous inhibition of bone destruction may seem like a good idea to prevent osteoporosis, continuous inhibition may actually be a really bad idea.
In addition to providing structural support and organ protection, bone serves as a repository of calcium, phosphorus, and mitogens (growth factors). Movement of these compounds into and out of bone is mediated by the two cell types (osteoclasts and osteoblasts) which in turn are modulated by numerous endocrines produced throughout the body.
Osteoblasts (the bone building cells) secrete collagen and other bone proteins creating a matrix onto which calcium and phosphorus crystallize (~ 90% of bone mass). The calcium to phosphorus ratio in bone is 2.5 to 1. In addition to calcium and phosphorus , various growth factors (mitogens) are also stored in the bone. Osteoblasts arise from osteoprogenitor cells located in the bone marrow and periosteum. Osteoprogenitors are induced to differentiate under the influence of growth factors, including the bone morphogenic proteins, fibroblast growth factor, platelet-derived growth factor.
Osteoclasts (the bone dissolving cells) secrete both proteolytic and hydrolytic enzymes and hydrochloric acid that result in destruction of the bone’s protein matrix, which results in mobilization of calcium, phosphorus, and bone resident growth factors into the extracellular fluid. Osteoclasts arise through the differentiation of macrophages. Osteoclasts are regulated by several hormones including PTH from the parathyroid gland, calcitonin from the thyroid gland, estrogen, vitamin D, and growth factor interleukin 6 (IL-6). Osteoclast population density is modulated by three molecules produced by osteoblasts - two that promote osteoclast development and one that suppresses osteoclast development. The two osteoclast promoter molecules are 1) macrophage colony-stimulating factor that binds to a receptor on macrophages inducing them to multiply and 2) RANKL (receptor activator of NF-kB ligand) that binds to a different receptor (RANK receptor) inducing the macrophage to differentiate into an osteoclast. The molecule that inhibits osteoclast formation is osteoprotegerin (OPG), which blocks osteoclast formation by latching on to RANKL and blocking its function.
The population density levels and activity levels of Osteoclasts and Osteoblasts are mediated by numerous endocrines. Osteoclast / Osteoblast mediated release of Ca++ (calcium ions), phosphorus, and mitogens from bone into the extracellular fluid has a profound impact on many physiological processes.
A summary of some of the more important endocrine interactions with the bone microenvironment that result in systemically increased extracellular calcium levels are listed below:
Increasing Vitamin D (1,25D) increases extracellular Ca++
Decreasing estrogen increases extracellular Ca++
Decreasing testosterone increases extracellular Ca++
Increasing prostaglandins increases extracellular Ca++
Decreasing growth hormones (GH, IGF, BMP) increases extracellular Ca++
Increasing parathyroid hormone (PTH) increases extracellular Ca++
Decreasing calcitonin increases extracellular Ca++
Increasing Vitamin A / Retinoids increases extracellular Ca++
Increasing Lithium increases extracellular Ca++
Calcium: Calcium is the fifth most abundant element by mass in the earth’s crust and fifth most abundant dissolved ion in sea water. It is especially important at a sub cellular level where movement of the calcium ion Ca++ into and out of the cytoplasm functions as a signal for many cellular processes. The average adult human body contains 1.3 kg of calcium of which 99% is contained in bones and teeth, 1% in cells of soft tissue, and 0.15% in the extracellular fluid. Intracellular cytosolic concentrations of Ca++ are kept low relative to extracellular concentrations. This concentration gradient drives Ca++ into the cell when Ca++ channels transiently open, which in turn activates Ca++ responsive proteins. Ca++ is required for function of nerves, muscles, secretions by cells, and cell growth and division pathways (population density management pathways).
Phosphorus: is involved in numerous physiological processes including transport of cellular energy via addition of a phosphorus atom to adenosine diphosphate (ADP) to make the energy rich molecule adenosine triphosphate (ATP). Phosphorus is important for key regulatory events such as phosphorylation (addition of a phosphorus atom to a molecule) and phospholipids are the main structural components of cellular membranes. Phosphorus is also used in maintenance of extracellular / intracellular ion concentration gradients via transmembrane ATPase pumps.
Growth factors: Growth factors were first discovered when scientists found that cells in culture would not grow and divide without growth factors. Today, more than 50 growth factors are known and are used in different combinations by the 210 different cell types in the body to maintain population density (i.e. replace dead cells, repair injury, increase select cell population counts as required by the body, etc…). Growth factors that are liberated by osteoclast activity include platelet-derived growth factors (PDGF), fibroblast growth factors (FGF), insulin like growth factors (IGFs) I and II, transforming growth factor-beta (TGF-beta), endothelin 1 (ET-1), urokinase type plasminogen activators, bone morphogenetic protein (BMP), and others. Growth factors (mitogens) released from bone play an important role in both bone rebuilding as well as population density management of various cell types throughout the body. PDGF and FGF stimulate progression of many cell types through the early part of the G-1 Phase of the cell division cycle and IGF-1 and IGF-2 are promote cell progression through the later part of the G-1 Phase. Released bone morphogenetic protein (BMP) is used to rebuild bone as well as maintain other tissues (cartilage, muscle, blood vessels) and organs (kidneys).
Growth Factors are used in the Population Density Management System of a multicellular organism. Cells posses growth factor receptors on their surface. Growth factors that bind to those receptors stimulate a cell to grow and divide (i.e. enter the Cell Division Cycle). Cells also produce molecules that are inhibitors of growth. Population density is a balance between stimulators and the inhibitors of growth.
Growth control pathways are in our DNA and determine things like how big we grow, the cell count of any one of the 210 different cell types relative to the others, and how tissue or organs are repaired after injury.
To provide a better understanding the complexity of the Cell Cycle Control System, the cell cycle control system for an Epidermal Growth Factor Receptor (EGFR) is shown below. The docking of a growth factor (mitogen) with the growth factor receptor results in its activation, which in turn results in the activation of a cascade of transduction molecules that transfer the signal to the nucleus, where transcription molecules are activated, that result in DNA expression (protein synthesis), that results in the production of cyclin and CDK proteins that drive the Cell Cycle Control System.
The levels of these Cell Cycle Control proteins rise and they form Cyclin/CDK complexes (graphic below). Complexes such as Cyclin D/ CDK and Cyclin E/CDK in turn function to hyperphosphorylate (add phosphor molecules to) the Rb block protein, removing it from the transcription factor it blocks, and allowing DNA transcription / production of proteins necessary for the cell to enter the next phase of the cell cycle.
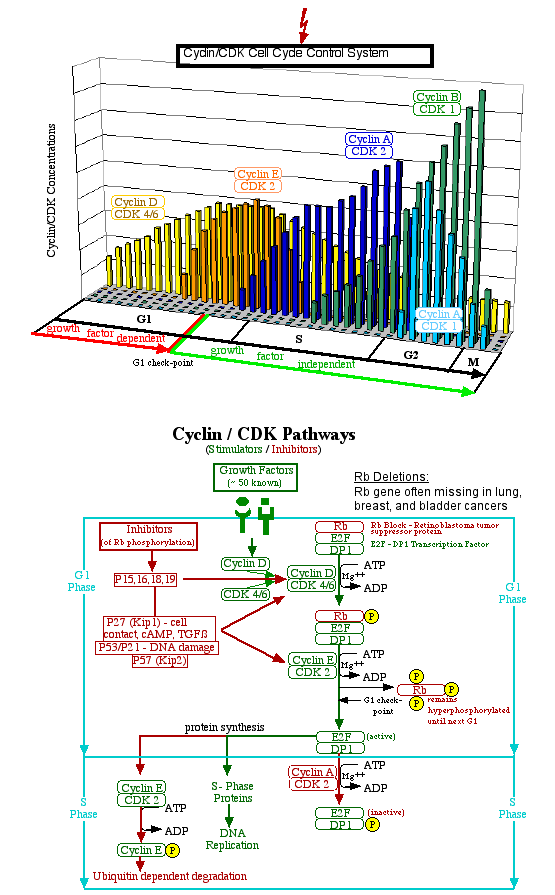
As an important side note, every time a cell divides, it accumulates on average three "genetic accidents" (mutations) from DNA replication errors. If those mutations occur in DNA expression related to growth control pathways, such as overexpression of growth factor receptors, receptors/ transduction molecules/ transcription molecules that are malformed to be "always active", or mutations that result in deletion of inhibitory proteins such as the Rb block protein, they can result in a cell that is programmed to relentlessly grow and divide, or what we call cancer.
That cancer cell will then relentlessly grow and divide into 2 cells, then 4, then 8, and so on, in what is called a first order logarithmic progression. In 30 population doublings it will have reached 1 billion cells (or a 1 cc clinically detectable mass). In 10 more population doublings it will have reached 1 trillion cells (or a 1 liter mass) which is lethal burden in a human.
Deposits to / Withdrawals from the Bone Pantry:
Parathyroid Hormone (PTH): The primary regulatory hormone responsible for increasing serum concentrations, by release of calcium from bone (bone resorption), is parathyroid hormone (PTH). When calcium sensors in the parathyroid gland detect low serum calcium concentrations, production of PTH is upregulated. PTH interacts with its receptor on osteoblasts to upregulate production of RANKL, which upregulates macrophage differentiation into osteoclasts. Additionally, PTH increases calcium reabsorption by the renal tubules (kidney) and stimulates conversion of vitamin D to its active form (calcitriol or D3).
Calcitonin: The primary regulatory hormone responsible for decreasing serum concentrations of calcium by inhibiting release of calcium from bone is calcitonin, which is produced by the parafollicular cells of the thyroid. High serum calcium concentrations result in upregulated production of calcitonin. Calcitonin receptors have been found in osteoclasts and osteoblasts and calcitonin result in the loss of the ruffled osteoclast border responsible for resorption of bone. Calcitonin also increases renal excretion of calcium by decreasing reabsorption by the kidneys and evidence exists that it reduces absorption of calcium in the gastrointestinal tract.
Estrogen has a “triple whammy” ( Rosen C J, “Restoring Aging Bones”, Scientific American, March 2003) effect in inhibiting osteoclast activity by binding to osteoblasts and 1) increasing their output of OPG and 2) suppressing their RANKL production. In addition, estrogen appears to prolong lives of osteoblasts while simultaneously 3) promoting osteoclast apoptosis. As estrogen levels drop after menopause, these “brakes” on osteoclast inhibition are removed, tipping the balance in favor of osteoclast dominated bone destruction which results in osteoporosis.
Androgens such as Testosterone also have an inhibitory effect on bone resorption, and studies suggest that this occurs through local aromatization of androgens into estrogen, however direct androgen interactions with androgen receptors related to bone remodeling have been observed in animal models.
Prostaglandins are autocrine and paracrine hormones produced in many places throughout the body. Prostaglandins have a wide array of effects, including involvement in the inflammatory part of an immune response to injury / cell death and antigens / allergens. Prostaglandins exhibit PTH-like (parathyroid hormone) effects that result in calcium / phosphorous/ mitogen mobilization from the bone, which is particularly useful in healing (population density restoration) in response to injury / cell death.
Vitamin D is a steroid-like chemical that promotes osteoclast activity by binding to vitamin D receptors (VDR) in osteoblasts and upregulating expression of RANKL, which upregulates macrophage differentiation into osteoclasts, which upregulate release of calcium ions, phosphorous, and growth factors from bone into the extracellular fluid. Vitamin D also enhances intestinal absorption of calcium and enhances renal retention of calcium. Its particular significance is discussed further below.
Skin - Bone - Ca++ Mediated Life
In context of the photosynthetic cyanobacterial DNA origins of life on earth, it is not surprising that vestiges of photo activated pathways that alter life functions can be found in humans today.
Skin exposure to sunlight (UVB) alters vitamin D levels, which in turn escalates extracellular calcium, phosphorus and mitogen levels, which in turn synchronizes a broad spectrum of day / night processes.
The active form of Vitamin D (1,25[OH]2D), also known as calcitriol or DHCC or 1,25 OHD or 1,25D, promotes osteoclast activity by binding to vitamin D receptors (VDR) in osteoblasts and upregulating expression of RANKL, which upregulates macrophage differentiation into osteoclasts (bone destroyers), that result in bone resorption (i.e. release of calcium, phosphorous, and growth factors stored in bone) . Vitamin D also activates absorption of calcium in the intestine and reabsorption of calcium by the kidney. Accordingly, the active form of vitamin D has a “triple whammy” effect on elevating extracellular calcium levels.
Normally, around 90% of the human requirement for vitamin D comes from exposure to sun. Skin is unique in that it is capable of manufacturing biologically active 1,25 D in the presence of UVB light from start to finish (unlike the “need regulated” conversion by the kidney). Full body exposure to UVB for 20 minutes in midday summer sun, in fair skinned people, can result in 10,000 IU of vitamin D being synthesized by the skin ( 25 times the recommended daily allowance of 400 IU). The effectively unregulated production of active 1,25 D by the skin would boost Ca++ levels by the three pathways previously discussed (i.e. increased release of calcium from bone, increased reabsorption of calcium by the kidneys, and increased absorption of calcium from the intestines) and the biological effect would last for a period of time commensurate with the amount of 1,25D synthesized and its half life ( 3 - 6 hours).
The other source of vitamin D synthesis is inside the body. The conversion of the inactive form of Vitamin D to the active form 1,25 D (calcitriol) involves two hydroxylations (addition of OH groups). The first hydroxylation is at the C-25 position and occurs in the liver through a cytochrome P-450 dependent enzyme and the second hydroxylation is at the C-1 position and occurs in the kidney. Parathyroid hormone (PTH) stimulates 1-hydroxylase and inhibits 25-hydroxylase. Calcitriol represses synthesis of 1-hydroxylase and enhances synthesis of 25-hydroxylase. Under normal conditions, low serum Ca++ levels increase PTH synthesis, which in turn increase conversion of vitamin D to its active form, which in turn elevates extracellular calcium levels by the three pathways disclosed above (i.e. increased release of calcium from bone, increased reabsorption of calcium by the kidneys, and increased absorption of calcium from the intestines). Elevated levels of the active form of vitamin D function to repress synthesis of 1-hydroxylase, which in turn functions to repress further conversion of vitamin D to its active form.
The Skin -Vitamin D mediated increase in daytime extracellular Ca++ levels result in enhanced nerve function, muscle function, and brain function, including consciousness and memory formation.
It should be noted that the concurrent decrease of another light mediated endocrine, melatonin (the sleep hormone), which drops with exposure to light and rises with exposure to darkness, likely creates a double whammy effect on daytime alertness. However, the focus of our discussion below is only on the Ca++ mediated enhancement of daytime functions.
The effects of increased daytime calcium levels on nerves is as follows:
a) nerve membrane depolarization
Increased extracellular Ca++ levels enhance nerve function. Since the active form of Vitamin D is synthesized during daytime, resulting in release of Ca++ from bone into extracellular fluid, nerve function is effectively heightened during daylight and reduced at night.
The basic science underlying neuron function were covered previously. This section covers more advanced neuron science.
Neurons contain ion channels that maintain an electrochemical concentration gradient between the outside of the cell and the inside of the cell. The resting membrane potential inside of a typical neuron is around -85 mV relative the outside of the cell (-30 mV to -100 mV depending on neuron type). The cell membrane acts as a capacitor, storing charge separated by the thickness of the membrane, and has a typical capacitance of about 1µ Farad per square centimeter. Changes in ion concentrations outside the cell versus inside the cell change the strength of the electric field across the cell membrane. Changes to the membrane potential are called “depolarizing” if they make the inside of the cell less negative or “hyperpolarizing” if they make the inside of the cell more negative. The term negative is relative, as it refers to an electrical potential differential between two points. If the inside is less positive than the outside, it is negative, relative to the outside of the nerve.
Electrical impulses that travel along the neuron are called action potentials and are transient perturbations in the membrane potential. Action potentials are conducted in a all-or-none manner and for an action potential to be generated the input signal must depolarize the neuron by more than its “threshold” membrane potential. As an example, for the -85 mV resting membrane potential neuron above, the threshold voltage is around -70 mV, meaning that the input signal must depolarize the membrane by at least 15 mV to generate a nerve impulse (i.e. action potential).
Changing the extracellular or intracellular concentrations of ions changes the resting membrane potential. Depolarizing concentrations (i.e. that make the inside of the cell less negative) bring the resting membrane potential closer to the threshold potential, and consequently the neuron requires a smaller input voltage to trigger an action potential. Polarizing concentrations (those that make the inside more negative) move the resting membrane potential farther away from the threshold potential and result in a larger input signal being required to trigger an action potential.
A traveling nerve impulse opens voltage gated Na+ (sodium) channels and K+ (potassium) channels, which allow Na+ to flow into the cell and K+ to flow out of the cell, passively along their respective electrochemical concentration gradients. Both the Na+ channels and K+ channels are rapidly inactivated by a “ball and chain” amino acid complex that rapidly plugs the respective channels. Potassium (K) is the most significant ion in impulse transmission because of the large disparity between the extracellular and intracellular concentrations. Typical extracellular concentrations potassium and sodium are about 3 mM of K+ and 117 mM of Na+ and the typical intracellular concentrations are about 90 mM of K+ and 30 mM of Na+ . The 30 fold concentration gradient disparity of K+ ( i.e. 90 / 3) overwhelms the 4 fold gradient disparity of Na+ (i.e. 117 / 30).
The resting (equilibrium or E) membrane potential for a given ion concentration can be calculated using the Nernst equation:
Ek = (RT/zF)(ln(Co / Ci))
where:
Ek is the equilibrium (or resting) membrane potential for the ion
R is the gas constant (8.31 joules/mole/ oK)
T is the absolute temperature (Kelvin = 273 +oC)
z is the valence of the ion (e.g. + 1 for potassium, + 2 for calcium)
F is the Faraday constant (amount of charge on a mole of ions, 96,500 coulombs/mole)
Co is the outside (extracellular) concentration of the ion (in mM)
Ci is the inside (intracellular) concentration of the ion (in mM), and
ln is logarithm to the base e
As an example, at room temperature (20oC = 293 oK) and for potassium (K):
RT/zF = (8.31)(293) / (+1)(96,500) = .02523 V = 25 mV
and for concentrations of 3 mM outside the cell and 90 mM inside the cell:
Ek = (25 mV)(ln (Ko / Ki)) = (25 mV)(ln 3/90) = (25 mV)(-3.4) = -85 mV
The effect of elevating extracellular concentrations of positive ions can be seen from the Nernst equation. Increasing extracellular concentration of the positive ion K+ results in a more positive resting membrane potential, which is by definition depolarizing, and brings the resting membrane potential closer to the threshold potential. This means a smaller input signal voltage is required to trigger the “all-or-none” action potential.
As an example, as extracellular concentrations of K+ are raised to 4 mM, the resting membrane potential becomes more positive (less negative):
Ek = (25 mV)(ln (4 / 90)) = (25 mV)(-3.11) = -78 mV
Using the -70 mV threshold voltage, the input voltage required to initiate an action potential is now only 8 mV versus 15 mV.
The actual resting membrane potential is a summation of all ions that are permeable. Calcium ions are permeable through the sodium - calcium exchanger (NCX).
From the Nernst equation, we can see that increasing extracellular concentrations of positive ions, relative to intracellular concentrations of positive ions, is a depolarizing change. Accordingly, elevating extracellular Ca++ levels relative to intracellular Ca++ levels is a depolarizing event that would lead to neuronal membrane depolarization (i.e. reducing the magnitude of the input signal required to initiate an action potential).
Neuronal intracellular Ca++ levels are kept low as calcium is a signaling molecule within a neuron (used for neurotransmitter release at the synapse). NCX in the plasma membrane and Calcium ATPase (PMCA) pumps in the synapse pump calcium out of the cytoplasm. Extracellular concentrations of Ca++ can range from 1 to 2 mM (Alberts B., et. al., 1994 p. 508). However, intracellular concentrations are kept very low and do not increase proportionately relative to extracellular increases. Studies of mammalian brain nerve cells showed that as extracellular concentration of Ca++ were raised from 1 mM to 2 mM, the intracellular concentrations only rose from 130 nM to 160 nM, respectively (Nachshen D. A., 1985) Accordingly, for a 100% increase in extracellular concentrations of Ca++, the intracellular concentrations only rise 25%.
From the above information we can approximate the amount of depolarization that would occur across the range of 1 mM to 2 mM of extracellular Ca++. Using the Nernst equation and the change in the ECa between the 2 nM and 1 nM levels would provide the amount of depolarization in mV that could be expected (per 1 mM) over this range (i.e. ECa @ 2 mM - ECa @ 1 mM = net change in resting membrane potential from a 1 mM change in extracellular Ca++ concentrations), or:
ê ECa per 1 mM increase in [Ca]o = ECa @ 2 mM - ECa @ 1 mM
For calcium, RT/zF = (8.31)(293) / (+2)(96,500) = 12.6 mV
and the ê ECa when extracellular Ca rises from 1 mM to 2 mM and the corresponding intracellular Ca levels rise from 130 nM to 160 nM:
= (12.6 mV )(ln (2 / .000160)) - (12.6 mV)(ln (1 / .000130))
= (12.6 mV) (9.43) - (12.6 mV)(8.948)
= + 6.12 mV
Accordingly, the increase in extracellular Ca2+ concentrations from 1 mM to 2 mM would make the resting membrane potential more positive by around 6 mV. In our previous example, this would reduce the resting membrane potential from -85 mV to -79 mV, which in turn would reduce the amount of input stimulus required to trigger a nerve impulse from 15 mV to 9 mV.
This nerve hypersensitization via neuronal membrane depolarization disclosed above is the first mechanism by which rising extracellular calcium ion concentrations affect the nervous system.
b) calcium mediated neurotransmitter release
The second mechanism is calcium mediated neurotransmitter release, via the voltage gated calcium channels.
As a nerve impulse reaches the synapses at the end of the nerve cell, it results in the release of chemicals called neurotransmitters, which in turn trigger a nerve impulse in the next cell in the transmission path. The electrical impulse causes voltage gated Ca++ channels to open which allows an inrush of Ca++ to enter the pre synaptic cell, along its electrochemical concentration gradient. Neurotransmitter is stored in vesicles at the synapse and Ca++ causes the vesicles to fuse with the cell membrane, releasing the neurotransmitter by exocytosis into the synaptic cleft. The neurotransmitter binds to and opens transmitter-gated ion channels on the post synaptic cell, which triggers a depolarization in the post synaptic cell, triggering an action potential if sufficient depolarization occurs. The extent of the depolarization of the post synaptic cell is graded according to how much neurotransmitter is released at the synapse and how long it persists there (Alberts B., et. al., 1994, p. 536).
For a 100% increase in extracellular concentrations of Ca++, the intracellular concentrations only rise 25%. As extracellular Ca++ levels increase from 1 mM to 2 mM, not only does the absolute amount of molecules available to rush in through the voltage gated channels double, but the concentration gradient (i.e. the driving force for the inrush) increases 63% from being 7,672 times greater on the outside at 1 mM ( i.e. 1 mM / 130 nM) to being 12,500 times greater on the outside at 2 mM ( i.e. 2 mM / 160 nM). Accordingly, the much larger amount of Ca++ entering the pre synaptic cell during the transient period when the voltage gated channels are open would result in much greater release of neurotransmitter.
Since depolarization of the post synaptic cell is graded and related to the amount of neurotransmitter released, as previously discussed, the effect of rising extracellular Ca++ levels would also be “ hypersensitization of synaptic gap transmission” via greatly upregulated neurotransmitter release from the pre synaptic cell combined with the neuronal membrane hypersensitization in the post synaptic cell (i.e. via the depolarization per the Nernst equation). Accordingly, rising extracellular calcium concentrations would have a direct “double whammy” effect on enhancing nerve function.
Extracellular Ca++ levels also have a direct effect on muscle tissue. Elevated extracellular calcium levels would increase muscle contraction by two pathways.
a) neuromuscular neurotransmitter release
The first relates to nerves and the neuromuscular junction. Muscle contraction is triggered by a nerve impulse traveling down a neuron which is then converted to a release of the neurotransmitter acetylcholine at the synapses where the neuron meets the muscle. Enhanced neurotransmitter release results when extracellular Ca++ levels are high, by the voltage gated Ca++ channel pathways previously disclosed above. Accordingly, more acetylcholine is released at the neuromuscular junction, causing a greater post synaptic depolarization.
b) sarcoplasmic reticulum calcium release channels
The second pathway relates to extracellular Ca++ concentration’s direct effect on muscle contraction. The release of the neurotransmitter acetylcholine described above causes the muscle to depolarize via neurotransmitter gated channels. The depolarization spreads along the muscle surface and the T-tubules that run along the surface of the muscle fibers. The depolarization opens voltage gated Ca++ channels in the T-tubule surface that allows Ca++ from the extracellular fluid in the T-tubule to enter the the sarcoplasmic reticulum. The inrush of Ca++ into the sarcoplasmic reticulum activates the “sarcoplasmic reticulum calcium release channels” (SRCaRCs), which in turn release Ca++ into the fluid around the myofibrils. The released Ca++ allows the muscle to contract by removing the tropomyosin block between actin and myosin, triggering cross-bridge formation by enabling myosin to bind to actin.
With an increase in the extracellular calcium concentration, there will be a larger release of Ca++ from the T-tubules, which in turn will activate more SRCaRCs and the release of more Ca++ onto the myofibrils, which in turn will cause greater cross-bridge formation and muscle contraction.
The skin - bone - Ca++ mediated daytime elevations in Ca++ levels enhance sensory and motor neuron function throughout the body.
The Calcium (Ca++) Wave, Consciousness and Memory Formation
The daytime enhancement in basic brain neuron activity would be in part attributable to the a) neuronal membrane depolarization and b) enhanced neurotransmitter release mediated by elevated Ca++ as previously discussed.
However, a very powerful effect on consciousness and memory formation would come from the elevated Ca++ levels affect on enhancing the propagation of the calcium wave in the brain. Propagation of the calcium wave in the brain has been associated with consciousness and memory formation.
The calcium wave is driven by a propagating chain reaction between a certain type of neuron ( glutamate activated, NMDA channel gated) and a certain type of glial cell (astrocyte).
Memory consolidation depends on astrocyte metabolism (Gibbs et. al., 2008). Astrocyte release of glutamate coupled with post synaptic depolarization leads to long term potentiation (Perea et. al., 2007). There is also evidence that “the informational content of perceptual conscious processes is embodied in astrocytic calcium waves” (Periera et. al. 2009).
Neurons make up only 10% of the brain and glial cells make up 90% of the brain. Astrocytes are the most numerous glial cells. Astrocytes are star shaped cells that make up one half of brain tissue volume. Astrocytes contact neuronal soma (cell body), dendrites, and presynaptic terminals of nerves. The dendritic tree of a neuron requires numerous astrocytes for complete coverage and conversely 300 - 600 dendrites are located within an astrocyte (Halassa et. al., 2007). The average cortical astrocyte also enwraps 4 neuronal cell bodies.
The contribution of Glial cells to intelligence and brain function is known, but has yet to be fully characterized. Empirical evidence strongly supports the correlation between higher glial to nerve cell ratios and intelligence. Moving up the evolutionary ladder (Koob, 2009): Leech -one glial cell per 30 neurons or 3% glial content, fruit fly: 20% glial, mice and rats 60%, Chimpanzee 80%, human 90%.
Ca++ Mediated NMDA ion channel activation
Glutamate is the main excitory neurotransmitter in the mammalian central nervous system. In the hippocampus, which is involved in the consolidation of short term to long term memory, there is a subclass of glutamate gated ion channels, known as NMDA receptors.
The NMDA channels are doubly gated, requiring that both glutamate to be bound to the receptor and the nerve membrane to be strongly depolarized (Alberts B., et. al., 1994, p 545). The second condition is required to release Mg++ that normally blocks the resting channel. Mg++ block removal combined with glutamate bound to the receptor allows Ca++ to enter the post synaptic cell. The increased Ca++ in the post synaptic cell produces a retrograde signal that produces a lasting change in the presynaptic cell that allows the presynaptic cell to release a greater amount of glutamate when subsequently activated.
Escalation in extracellular Ca++ functions to depolarize nerves by the Nernst equation as previously discussed. The depolarization in turn enhances removal of the Mg++ block, allowing glutamate mediated influx of Ca++ into the nerve cells.
Nerves, astrocytes, and the calcium wave
In an activated astrocyte cell, inositol trisphosphate (IP3) pathways are activated inducing the release of calcium ions from internal stores (mitochondria and endoplasmic reticulum) which in turn result in the astrocyte releasing Glutamate (Glu) extracellularly, which in turn binds to and activates receptors on post synaptic neurons that allow Ca++ inflow into the neuron.
A mechanistic explanation for the propagation of the calcium wave is not well elucidated in literature. Accordingly, author presents a mechanistic outline below of what has not been elucidated, but should be inferred from available studies.
Author would like to note a significant implication from the elucidation is that these specialized brain neurons can conduct signals in two ways instead of just one: one by the traditional Na+ / K+ driven membrane depolarization route and the second by Ca++ intra neuron propagation. Although each of the two transmission modes is antagonistic to the other, the combination of both being activated would effectively provide a third state that appears may be responsible for long term memory formation.
Starting with the above glutamate activated inrush of Ca++ into a neuron, the inrush of calcium would be expected to propagate along the concentration gradient differential toward the synapse.
From the Nernst equation, we know the intracellular Ca++ inrush is a hyperpolarizing event. As an example, if the inrush transiently doubled intracellular Ca++ levels to 320 nM from 160 nM:
ê ECa = (12.6 mV )(ln (2 / .000320)) - (12.6 mV)(ln (2 / .000160))
= (12.6 mV) (8.74) - (12.6 mV)(9.43) = (110.1 mV) - (118.8 mV) = - 8.7 mV
Accordingly, the transient increase in intracellular Ca++ concentration would make the resting membrane potential more negative (i.e. less positive) by around 9 mV (from -85 mV to - 94 mV), boosting the amount of input stimulus required to trigger a nerve impulse from 15 mV to 24 mV for the traditional Na+ / K+ driven depolarization.
Because the two transmission modes are antagonistic, it would require a much higher input voltage for a double mode firing to occur simultaneously. While not normal, a double firing, or this "third state" is not impossible, and could be expected in heightened excitory situations. It is possible this third state may involved in enhanced memory formation in heightened excitory situations.
In the normal Ca++ only firing mode, the inrush of calcium would propagate along the concentration gradient differential toward the synapse. Once the Ca++ reached the synapse it would fuse the vesicles containing neurotransmitter to the membrane, releasing the neurotransmitter (glutamate). The end result is effectively the same as that of a Na+ / K+ driven action potential opening voltage gated Ca++ channels at the synapse, fusing vesicles, and releasing neurotransmitter. The Ca++ would then pumped out of the cell by PMCA pumps at the synapse. A double firing could be expected to release a much larger amount of neurotransmitter and Ca++ into the synaptic cleft.
Released glutamate in turn triggers the next astrocyte in line, as well as directly activates any nearby post synaptic NMDA gated neurons, propagating the calcium wave in a process that is repeated until the end of the propagation path.
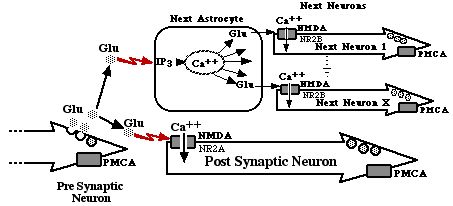
While the above physiological elucidation deals with how the propagating calcium wave is created, it does not deal with the observable electromagnetic phenomena associated with consciousness and memory formation. An elucidation of that is next.
The Calcium Wave Driven Electromagnetics of Consciousness and Memory
There is a clear electric / electromagnetic connection to consciousness and memory formation that has not been elucidated in literature, and author will attempt to elucidate it below:
Simplistically, the propagation of the calcium wave is likely what hosts the electromagnetic phenomenon responsible for consciousness and memory formation. Mechanistically, a traveling ion wave could act as an “electron broom”, unidirectionally moving ambient unbound electrons, which is what an electrical current is, and what is measured in most brain studies.
An electric current is defined as a flow of electrons. A traveling perturbation in ion concentration gradients is not a true electrical current per se. However, true electrical currents were measured by Dr. Becker ( Robert O. Becker MD and Gary Selden, “The Body Electric - Electromagnetism and the Foundation of Life“, © 1985, First Quill Edition.)
Dr. Becker found a negative electrical potential at the front of the head versus a positive potential at the back of the head, which suggested a direct current flow from the back of the head to the front of the head. Negative potentials in the brain’s frontal area and at the periphery of the nervous system were associated with wakefulness, sensory stimuli, and muscle movements. The more activity the greater the negative potentials were. A shift toward the positive occurred during rest and even more so during sleep.
An Electroencephalogram (EEG) records the underlying voltage fluctuations over various parts of the head. The frequency of these brain waves are correlated with states of consciousness. Beta waves (~ 15 - 30 cycles per second) are observed during daytime consciousness, in contrast to Delta waves ( ~ 1 - 3 cycles per second) which are observed during deep sleep.
The source of the electric current has never been elucidated. However, from physics we know that an electric field extends outward from electrically charged particles such as ions, which can in turn interact with other nearby electrically charged particles such as electrons (i.e. attract or repel them). A traveling ion perturbation wave could attract or repel electrons directionally along the path of travel, which would in turn create the electrical current measured by Dr. Becker.
The traveling electric current is required for consciousness and memory formation. The most compelling data comes from inhibiting this flow of electrons, which result in loss of consciousness and memory formation.
A strong magnetic field oriented at right angles to an electric current magnetically “clamps” or stops its flow. Dr. Becker (Becker et. al., 1985, p. 238) found he could anesthetize animals using this process, just as well as with chemicals, and found EEG recordings of magnetic and chemical anesthesia were identical. The absence of consciousness and memory formation during anesthesia induced by magnetic clamping of the electrical current flow confirms the required involvement of the moving electrical current in both consciousness and memory formation.
Alternatively, an opposing current source can be used to stop the flow of current. When Dr. Becker (Becker et. al. 1985, p111) passed a minute current from front to back through the head to cancel out the internal current, the animal fell unconscious.
Electrons have a magnetic field associated with them (because of their spin).
A traveling direct electric current is accompanied by a traveling magnetic field, which is perpendicular to the direction of the electron flow, and proportional to the strength of the current. A traveling magnetic field can interact with its surroundings in one of 3 ways: 1) induce a current flow in a conductor that is perpendicular to the direction of motion of the magnetic field, 2) deflect electrons flowing in a conductor so as to flow perpendicular to the direction of current flow (Hall effect), and 3) magnetize, or alter the magnetic orientation of, magnetic material in its path.
The human brain produces steady DC magnetic fields one billionth the strength of earth’s field of about one-half gauss (Becker et. al. 1985, p240). The mineral magnetite (Fe3O4) is known to be precipitated biochemically by bacteria, mollusks, arthropods, chordates, fish, animals, and humans and appears in the fossil record extending back to the Precambrian time (Kirschvink et. al., 1992). Kirschvink et. al. used high resolution transmission electron microscopy to estimate the presence of a minimum of 5 million single-domain crystals per gram of human brain tissue, with 100 million+ crystals per gram for pia and dura. The crystals are in clumps of 50 - 100 particles. The crystal alignment was interpreted as a biological mechanism for maximizing the magnetic moment per particle, as the direction yields 3% higher saturation magnetization than do other directions.
Magnetite is known to be used in geomagnetic orientation, indicating it interacts with brain tissue, possibly through the tissue’s associated electric field.
Whether the brain’s electric currents, associated magnetic fields, and magnetite play a role in memory or brain functions other than geomagnetic orientation has yet to be studied. In electronics, electric currents, magnetic fields, and magnetizable materials are used to store and recall memory. As an example, in a video camera, light entering the camera is converted to a set of electrical pulses that are converted at the recording head into magnetic fields that in turn magnetize the magnetic material on the tape below. This creates a record of the electrical pulse pattern. In playback mode, the play head is passed over the tape an reconverts the magnetic pattern back into the electrical pulse pattern as originally seen.
In summary, the effect of skin-bone mediated daytime escalation of extracellular Ca++ on the calcium wave would be to: 1) elevate intracellular astrocyte Ca++ levels, hence enhancing the astrocyte intracellular Ca++ mediated glutamate release, 2) elevate extracellular Ca++ levels to depolarize neurons and remove the Mg++ block from NMDA receptors, allowing glutamate to open Ca++ channels. The elevated extracellular calcium would also 3) provide a greater influx of calcium into neurons when the glutamate mediated Ca++ channels were open, which in turn would 4) result in a greater release of neurotransmitter when the higher levels of Ca++ reached the synapse. The combinational effect would be enhancement of the calcium wave. The calcium wave in turn may be what drives the observed directional flow of electrical currents (and their associated magnetic fields) in the brain, which in turn are associated with consciousness and memory formation by pathways which have yet to be elucidated.
The abundance of magnetite crystals in the brain, combined with the presence of magnetic fields in the brain, the strength of which is proportional to the underlying electric currents, which in turn would be expected to be proportional to the strength of the underlying calcium wave, indicate a possible novel mechanism for memory formation via magnetization of the magnetite crystals.
Chapter 6
Gallery of the Unknown: Indeterminate / Unknown Life Forms
The indeterminate and unknown fossils in this chapter represent life forms created by the great explosion of life that never made it into earth's playbook of life.
Speculative Evolution is defined in Wikipedia as a genre of speculative fiction focused on hypothetical scenarios in the evolution of life. While the fossils in this section are not "fiction" and were created by genetic reassortment and not evolution, they do provide a glimpse of a world that might have been. They are a testament to the great power of the DNA reassortment process to create new life forms, as well as a testament of how quickly a new ecosystem emerges and whittles down this population into a small subset by survival of the strongest.
The fossil specimens in this section exhibit the same lack of post mortem degradation and deformation consistent with the " high energy death by limestone fossilization" as disclosed in Chapter 1. As such, they appear to capture the last instant in the life form's life. They include hardened calcium carbonate fossils, crystalline calcium carbonate fossils, and the resin embed fossils as observed in the identifiable life forms and shown in Chapter 1.
The Indeterminate, Unknown, or other life forms that have no clear antecedent lineage or no presence on earth today (collectively the Speculative Evolution Fossils) are the second largest group of fossils found, just behind the CSFF fossil group that hosted the grab bag DNA reassortment process that created them.
Had the carnivorous dinosaurs not appeared, some of these alternate life forms may have gone on to inhabit the earth and form an alternate ecosystem.
Photos of 20 such life forms were presented in 2020. We added another 20 in June of 2021. We plan on adding 10 more in 2022.
Photo Gallery of the Unknown
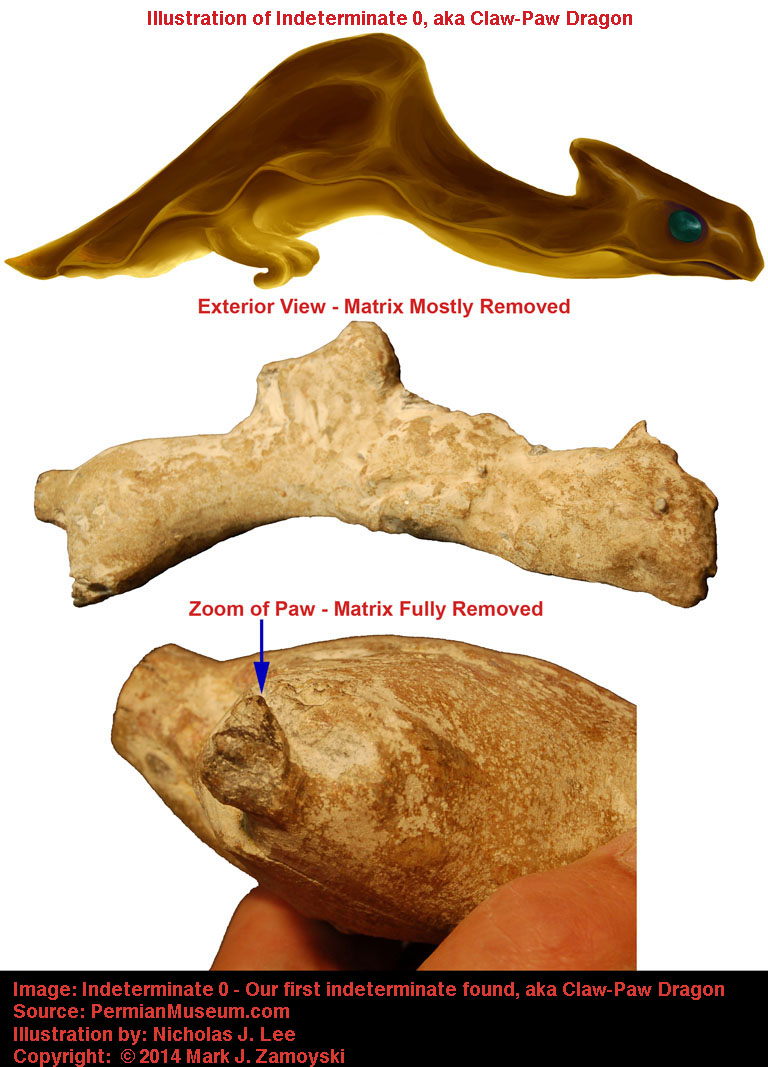


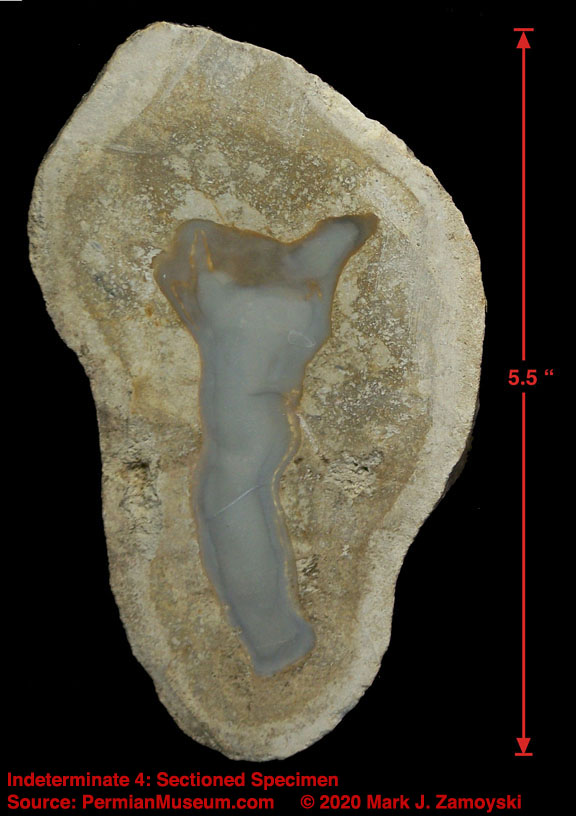

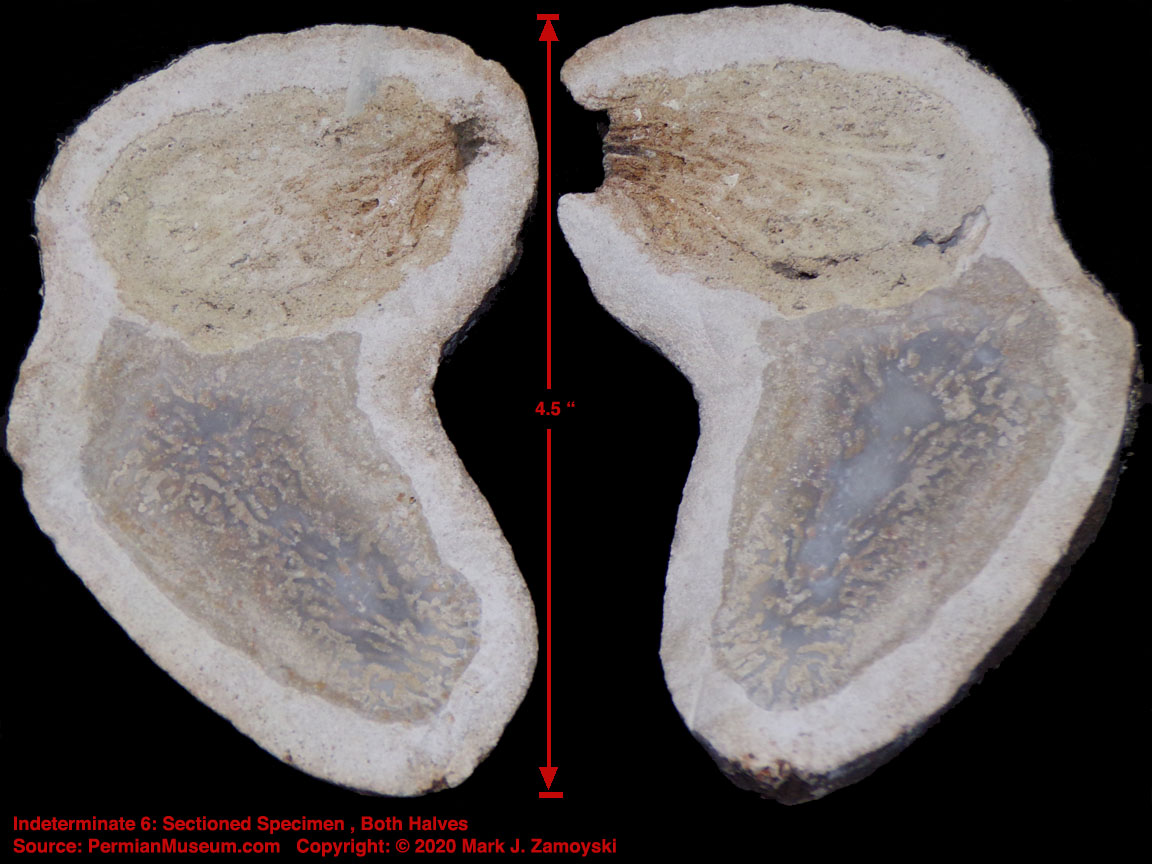





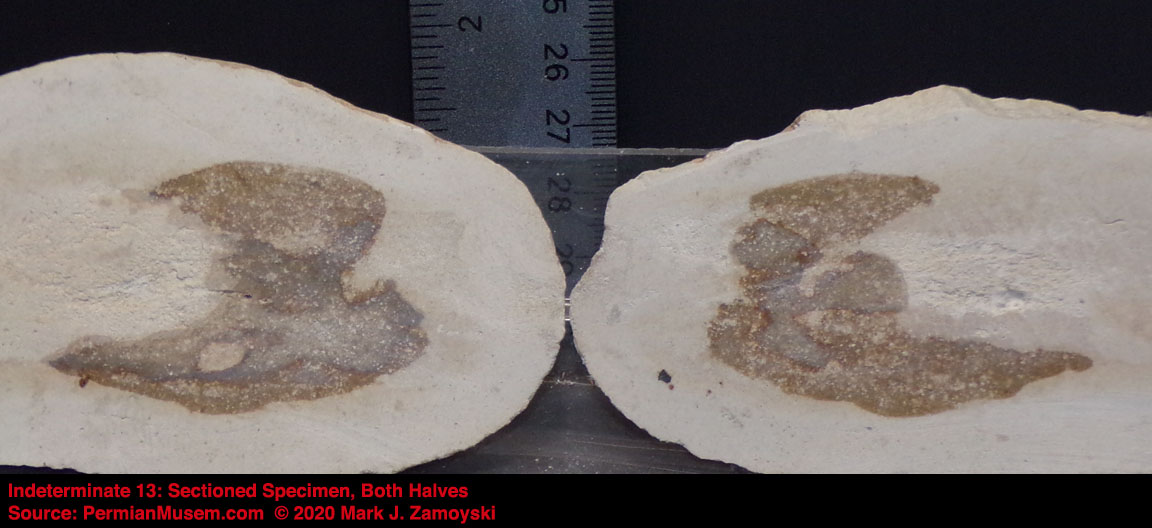
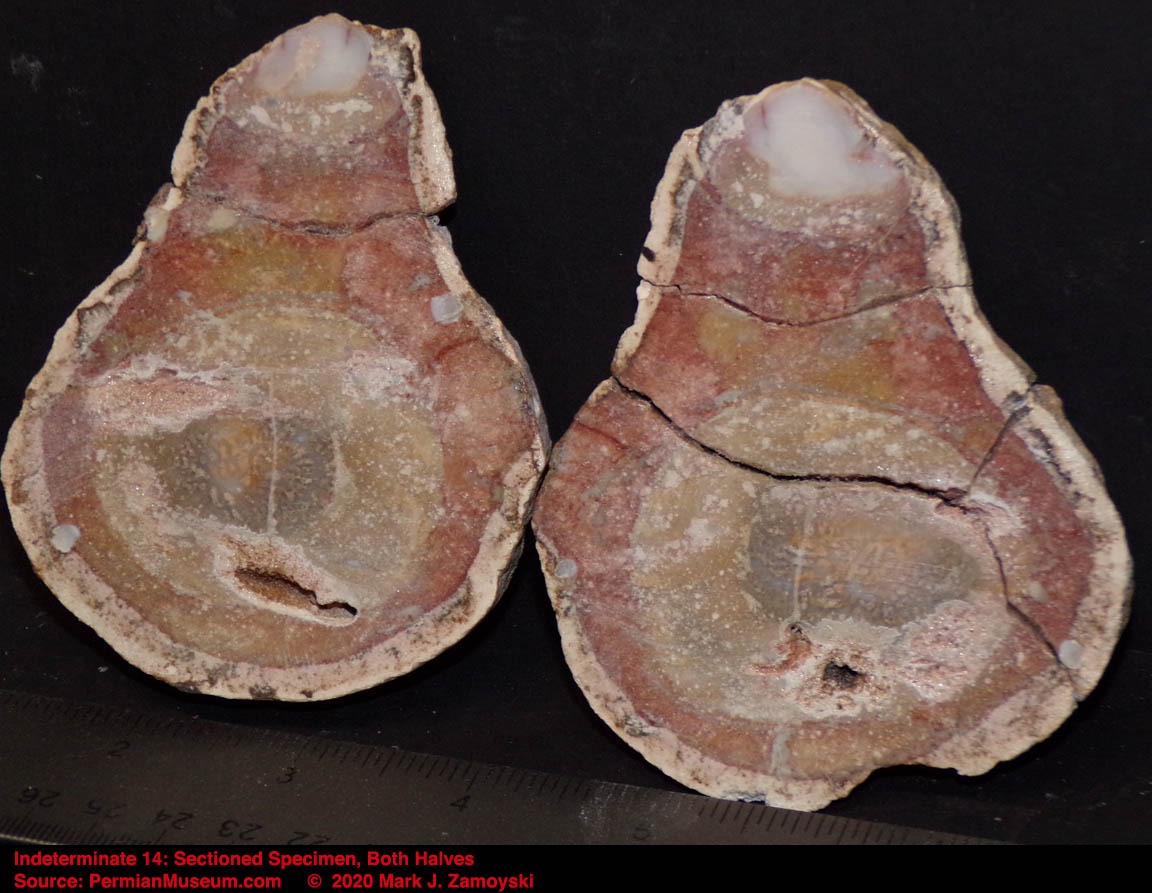

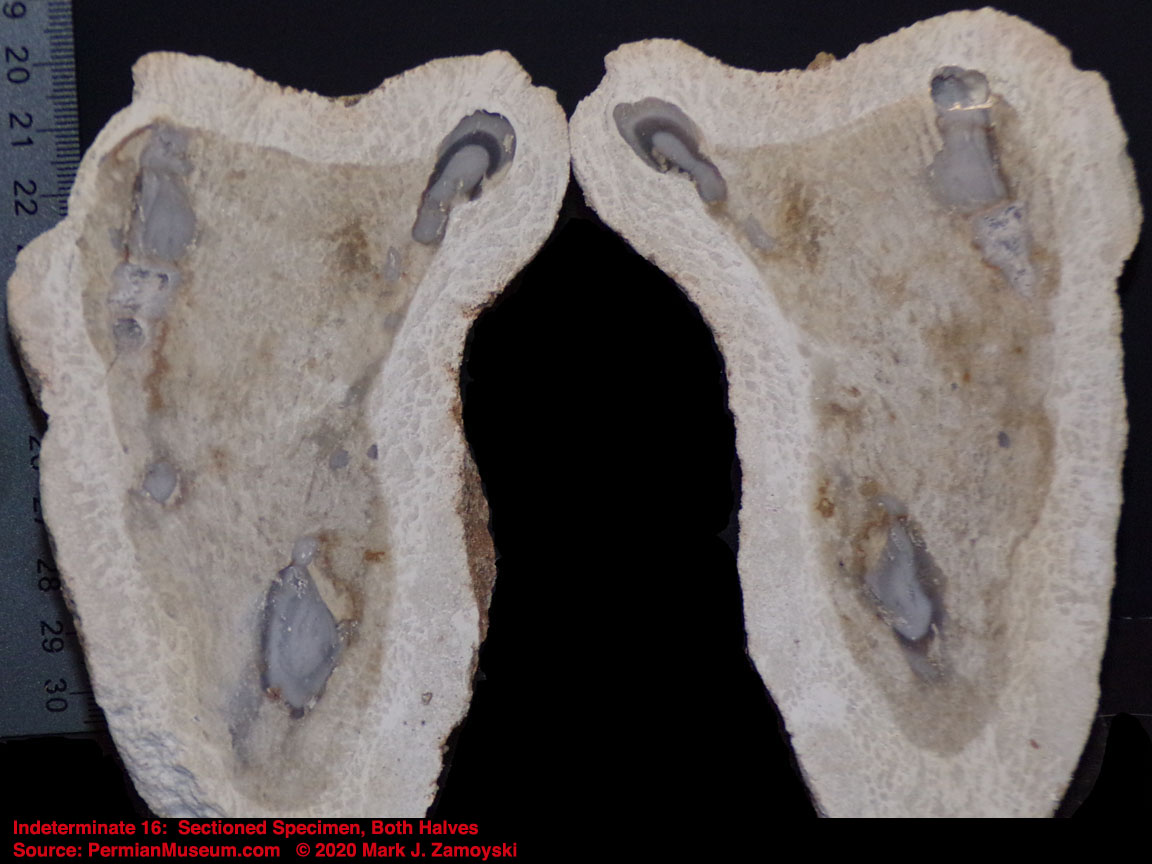


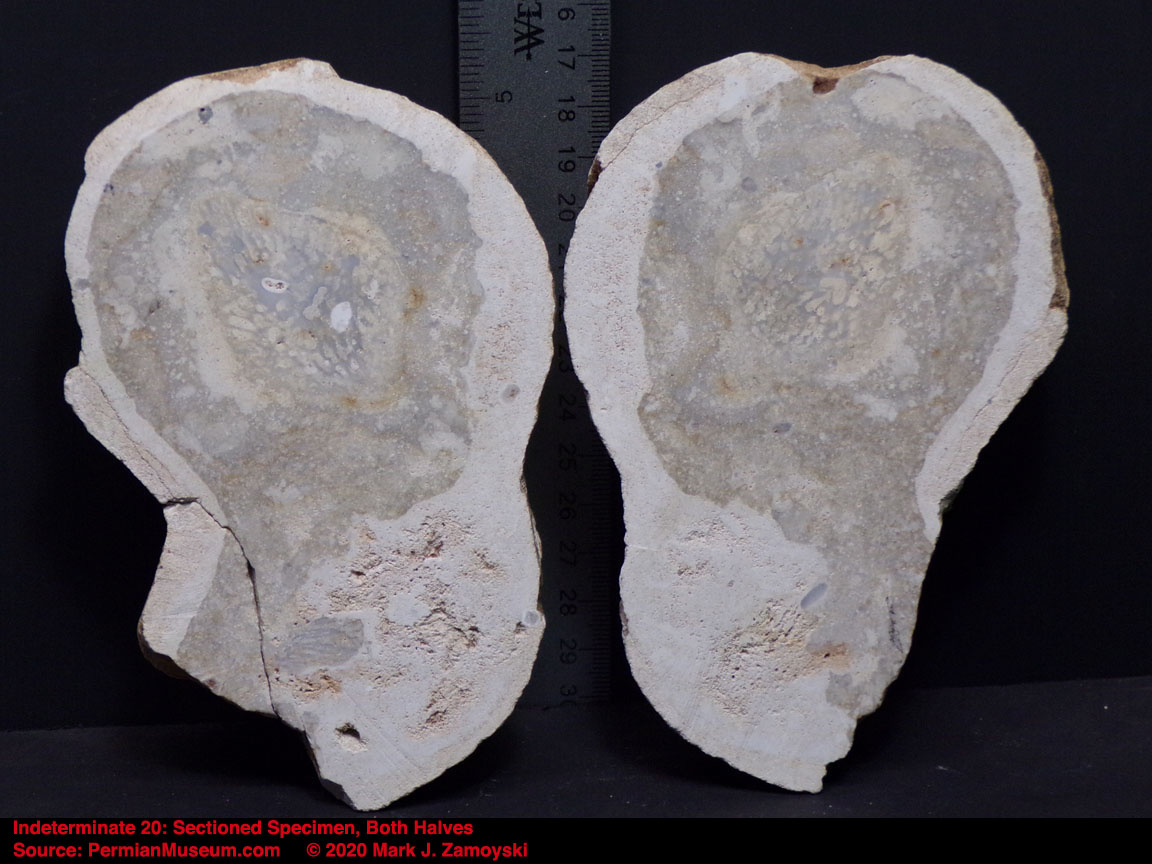

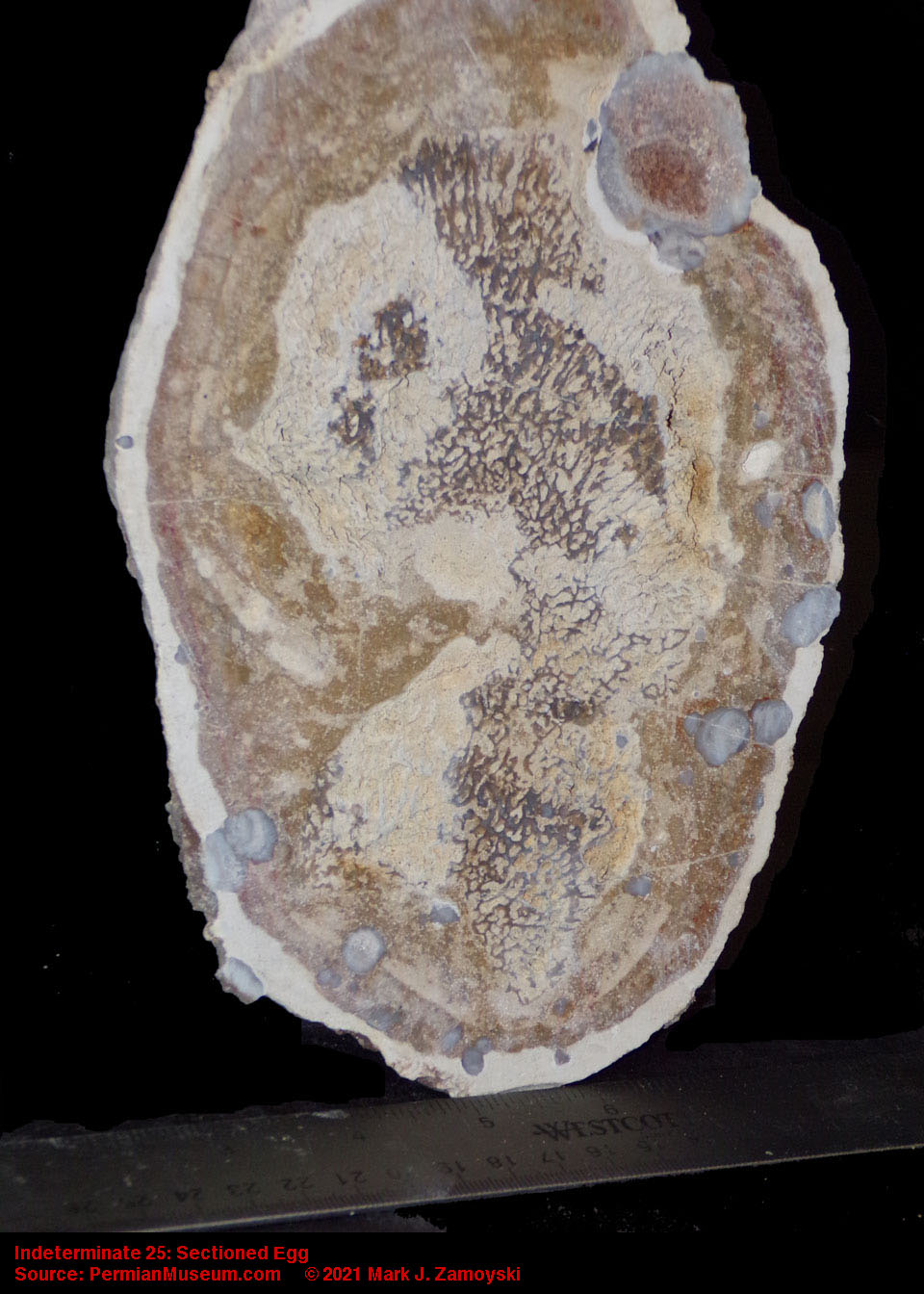









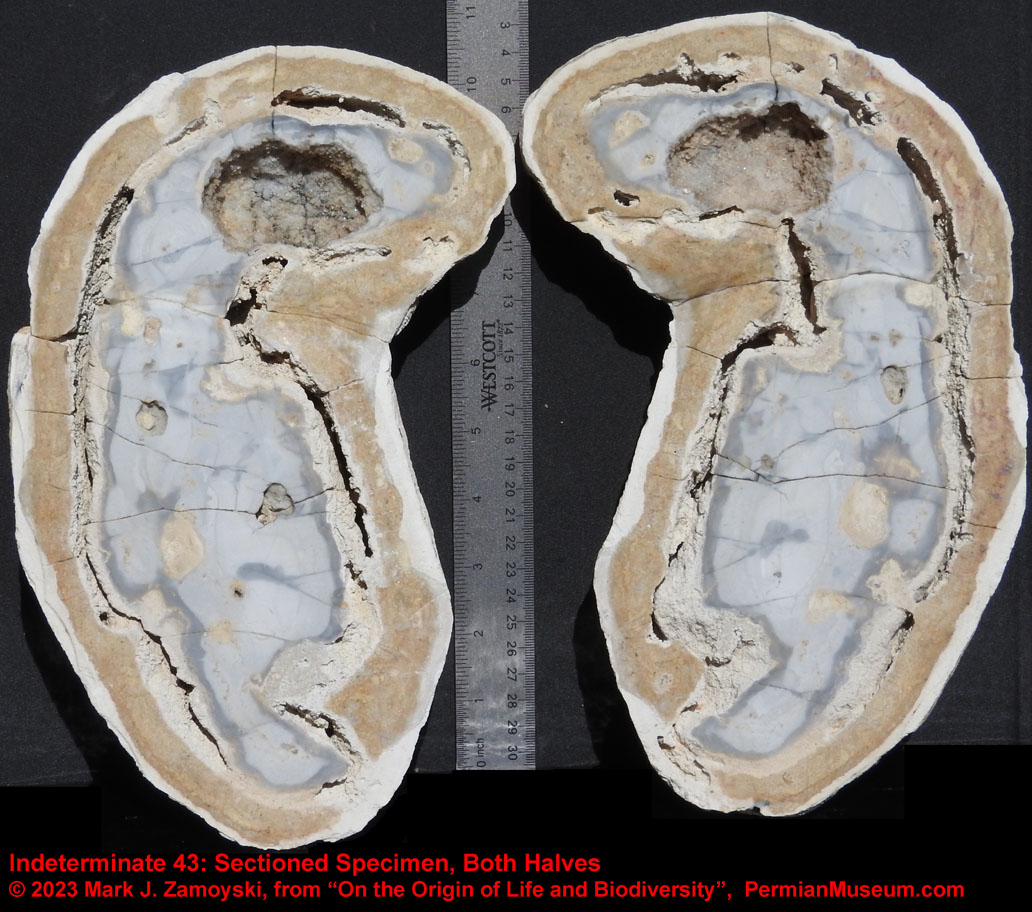
Chapter 7
Summary, Conclusions, and Looking Forward

Summary
The single celled prokaryotes that spawned life on earth first appear in the fossil record 3.8 Ba years ago, shortly after the peak of the asteroid impacts 3.9 Ba, and at around the same time the oceans formed 3.8 Ba.
They had a tough triple layer cell wall, can live two miles deep in soil at pressures of 5,000 PSI, can survive in anaerobic or aerobic environments, and can thrive near volcanic vents 3,500 feet below the ocean surface.
They have circular DNA, capable of generating around 1,000 proteins per million base pairs of DNA, versus humans which only crank out 7 proteins per million base pairs of DNA.
They also brought mitochondrial DNA to earth. Mitochondria is a cell’s power plant that stores energy from aerobic respiration (metabolism of glucose). Mitochondria is of prokaryotic origin: its DNA is separate from that of the nucleus, is circular (bacterial), and its nucleotide sequence analysis points back to early bacterial origins (rickettsia, rhizobacteria, and agrobacteria). Physiological life can be defined as a collection of concentration gradients. The presence of concentration gradients means life. The absence of concentration gradients means death. It takes energy to maintain concentration gradients. Without the prokaryotic mitochondrial DNA that arrived on earth some 3.8 Ba there would not be a sustainable energy source in cells and life as we know it today would not exist.
The prokaryotes also transformed earth into a habitable planet. Cyanobacteria were among the first prokaryotic cells to appear and can live in anaerobic or aerobic environments. They obtain their energy by photosynthesis, taking an electron from water and releasing oxygen as a waste product. They are believed to have created our oxygen atmosphere. Atmospheric oxygen began appearing around 2.5 Ba, and was close to current levels by 1.5 Ba, which is also when the unicellular eukaryotic explosion of life occurred.
Cyanobacteria also convert carbon dioxide into carbohydrates, providing a food source, and make the enzyme nitrogenase, which fixes nitrogen into a form that can be absorbed by plants and used in the synthesis of proteins and nucleic acids.
Prokaryotic cells took two paths in direct creation of DNA based life forms: 1) Self Replication and 2) Genetic Reassortment into new life forms.
The Self Replication route was so successful that today bacterial biomass on earth exceeds that of all plants and animals combined. One gram of soil contains 100 million to 1 billion bacterial cells. Coastal oceans contain 1,000,000 cells per ml. Should the earth ever be blown to pieces and hurled through space, the resulting pieces would effectively serve as seeds of life for new planets.
The Genetic Reassortment route is what we know as life on earth today. It has changed over time, from aquatic life, to the rise of the dinosaurs and winged flight, to the rise of mammals and primates. Prokaryotic DNA appears to also contain sequences that facilitate the DNA reassortment process -- such as that which codes for the class of single celled organisms known as the Calcium Secreting Filter Feeders, that build structures capable of hosting the DNA reassortment process (Chapter 3). The life forms crated by this genetic reassortment process (Chapter 4) were the explosion of life that introduced the dinosaurs, winged flight, and the rest of the reptiles, mammals and primates that formed the basis of life on earth today.
Conclusions
Based on the above, every time a meteorite or asteroid hits earth, there is the possibility of new DNA being introduced into the mix.
Every time there is a mass extinction event, the stage is set for a new set of life forms to emerge and form a new ecosystem.
Who or what created the prokaryotic seeds of life that arrived on earth some 4 Ba is still an enigma.
Over 4 billion years, no natural process on earth could create such a "seed of life". Instead, the genetic reassortment process took exactly the opposite route, devolving into a genetic debris field, coding for fragile life forms.
The only way a "seed of life" like Cyanobacteria could be created on earth is by artificial engineering of such a strand of DNA. That takes intelligence. That implies one possibility is that an extremely intelligent life form existed more than 4 billion years ago, and for some reason had the desire to create such a seed. Natural processes work by random chance, which pretty much makes it a mathematical impossibility that a Cyanobacteria's ~ 6 million base pair circular DNA could have been created by chance (i.e. a natural processes).
It is also mathematically impossible for natural processes to have created the fine tuned complexity inherent in our DNA. Depending on random chance is like randomly removing or adding parts to a finely tuned watch and expecting it to run better.
As an example, Growth Control or Population Density Management Pathways are a subset of Cell Signaling Pathways. A human has around 210 different cell types in the body and cells posses growth factor receptors on their surface (more than 50 are known). Cells also produce and release growth factors (mitogens) that bind to those receptors and stimulate a cell to grow and divide (aka enter the Cell Division Cycle). Cells also produce molecules that are inhibitors of growth. Population density is a balance between stimulators and the inhibitors of growth.
Growth control pathways are in our DNA and determine things like how big we grow, how tissue or organs are repaired after injury, the cell count of any one of the 210 different cell types relative to the others, etc....
To understand the mathematical impossibility, all one has to do is look at the complexity of the Cell Cycle Control System pathways for a single such receptor. The Cell Cycle Control System for an Epidermal Growth Factor Receptor (EGFR) is shown below. The docking of a growth factor (mitogen) with the growth factor receptor results in its activation, which in turn results in the activation of a cascade of transduction molecules that transfer the signal to the nucleus, where transcription molecules are activated, that result in DNA expression (protein synthesis), that results in the production of cyclin and CDK proteins that drive the Cell Cycle Control System.
The levels of these proteins rise and they form Cyclin/CDK complexes (graphic below). Complexes such as Cyclin D/ CDK and Cyclin E/CDK in turn function to hyperphosphorylate (add phosphor molecules to) the Rb block protein, removing it from the transcription factor it blocks, and allowing DNA transcription / production of proteins necessary for the cell to enter the next phase of the cell cycle.
As for the random chance aspect, every time a cell divides, it accumulates on average three "genetic accidents" (mutations) from DNA replication errors. If those mutations occur in DNA expression related to growth control pathways, such as overexpression of growth factor receptors, receptors/ transduction molecules/ transcription molecules that are malformed to be "always active", or mutations that result in deletion of inhibitory proteins such as the Rb block protein, they can result in a cell that is programmed to relentlessly grow and divide, or what we call cancer.
As for the mathematical impossibility aspect, you can pound a puddle of water containing amino acids and nucleotides with lightning for billions of years and you are not going to create something like the molecular pathway above. And the above is just one of hundreds of pathways.
Getting back to the historical record, the quantum leap in life forms spawned by the DNA reassortment process during the Permian Explosion of Life set the stage for life as we know it today. Dinosaurs had a great run, lasting ~ 180 million years, until 66 Ma when an asteroid impact caused the next mass extinction event.
The next 66 million years that followed saw the painfully slow evolutionary process transform the composition of life on earth once again.
Looking Forward
The K-T Extinction and the Rise of Humans
Unlike the end Permian extinction event that wiped out 96% of marine species, the Cretaceous-Tertiary (K-T) extinction event only wiped out 75%.
Growth control pathways became the determining factor in 1) what survived the asteroid impact as well as 2) what would become the new dominant life form on earth.
1) No tetrapods weighing more than 25 kilos (55 pounds) survived the global cooling caused by the asteroid impact.
2) Population density of glial cells (glial cell hypertrophy) determined the future direction of life on earth.
A single mutation or inherited trait in population density pathways, that is present in the first cell from which a life form arises (what we will call a birth mutation), can occasionally result in a greatly improved life form. A single mutation in the LRP5 gene results in a human with bone mineral density closer to that of a bear than a human. A mutation in the MSTN gene results in muscle hypertrophy, which basically means you look like a body builder without ever having to step into a gym. About 80% of an individual's height is determined by the DNA they inherited.
The dramatic climate change caused by the K-T asteroid was relatively short. The freezing temperatures reversed after 3 years (Brugger, Julia; Feulner, Georg; Petri Stefan (2016) "Baby, its cold outside: Climate model simulations of the effects of he asteroid impact at the end of the Cretaceous" ) Geopysical Research Letters. 44(1): 419-427) and returned to normal within decades (Whittle et. al., 2019, "Mass Extinction", Dryad Digital Repository).
Birth mutations, favorable breeding, including cross breeding, and selective advantage drove the painfully slow development of life, from antecedent lineages, after the K-T extinction event. Unlike the abrupt explosion of new life forms from genetic reassortment, the "evolutionary" process occurred in extremely small increments, over the next 63 million years.
The DNA of mammals and reptiles is 89% the same. Mammalian species began diversifying ~ 30 million years before the K-T extinction event.
The Paleocene period ( 66 - 56 Ma) that followed saw a tropical, greenhouse climate with forests worldwide and no polar ice. Global average temperatures were ~ 24 C (76F) or 10 C warmer than the 1951-1980 average of 14C (57F), and CO2 levels were around 500 ppm versus 400 ppm today.
The Eocene period ( 56 -34 Ma) that followed got even better, after 2,500 -4500 gigatons of carbon were released into the atmosphere, with the highest measured levels of CO2 reaching 4,000 ppm, and average temperatures rising by 5 - 8 degrees C (9 -14 F), which put the temperature around 15C ( 28 F) warmer than today. By the end of the Eocene, CO2 levels had decreased to around 800 ppm.
This climate is what hosted the rise of our forerunners. Primates are believed to have arisen from small terrestrial mammals and the first small monkey like life form (Haplorhini) appeared 63 Ma, or 3 million years after the K-T extinction event. Other small monkeys and apes appeared around 40 Ma. The great apes (gorilla, orangutan, chimpanzee, bonobo) appeared around 17 Ma. The Homo ("man" in Latin) genus first appeared ~ 2 Ma with Homo Erectus, which went on to spawn the Homo Neanderthalensis and Homo Sapiens (anatomically modern humans), which first appeared ~ 300,000 years ago. The earliest homo sapien fossils found so far date back to ~ 315,000 years ago.
Homo Sapiens dispersed from Africa in several waves, with the great "Southern Dispersal" believed to have started around 70 - 50,000 years ago, leading to the lasting colonization of Eurasia and Oceania by 50,000 years ago.
Humans (homo sapiens) share 98 % of the same DNA as chimpanzees, however, 90% of the cells in the brain of a human are glial cells, versus 80% for chimps. Glial cell, or more specifically Astrocyte cell hypertrophy, is a small population density mutation that made a big difference.
Glial cells are associated with intelligence. Astrocytes are the most numerous glial cells and are responsible for propagation of the calcium wave in the brain, which in turn hosts the measurable electromagnetic phenomenon underlying consciousness and memory formation, as was covered in Chapter 5. Neurons only make up 10% of the brain. The contribution of Glial cells to intelligence and brain function was also empirically presented in Chapter 5 (Koob, 2009) as the % of Glial content in various life forms:
- Leech: 3%
- Fruit Fly: 20%
- Mice and Rats: 60%
- Chimpanzee: 80%
- Human: 90%
That brings us to "intelligence", which seems to be the end product of 4 billion years of prokaryotic transformation of earth, followed by genetic reassortment and the explosion of multicellular eukaryotic life, followed by evolution and the selective advantage of Astrocyte cell hypertrophy.
So how does our intelligence (astrocyte cell hypertrophy) compare to the intelligence of whoever or whatever created the seeds of life that spawned us?
Would it be as described in the movie Men in Black , when an alien gave his description of humans as: "Barely conscious pond scum, convinced of their own superiority, living short, pointless lives."
There were geniuses like Einstein and Milankovitch that could visualize or conceptualize things that were not known or taught. They wrote new chapters in science, rather than just reciting chapters that had already been written. However, these are only two people out of billions.
Then there are the transient epiphanies, or moments of brilliance, we all have, where our minds make quantum leaps in intelligence and we figure things out that have long eluded us. Then comes the wave of humility, as we realize what our minds are capable of, and how infrequently they operate at that level.
So, what would an accurate assessment of human intelligence be? Let's take a look at some empirical data.
Case Study: Percentage of "Barely Conscious" Humans: 42%
On February 8, 2008, ~ 8 million people in California voted on Prop 91, providing invaluable data on what percentage of humans can be defined as "Barely Conscious".
The Official Voter Information Guide from the secretary of state contained the Pro and Con Arguments for an originally Democrat sponsored measure that was no longer needed. The Pro side simply said : "Prop 91 is NO LONGER NEEDED. Please VOTE NO. Voters passed Proposition 1A in 2006, accomplishing what Prop. 91 set out to do.
The "CON" side provided no argument, as the "PRO" side said it all.
The full text of the actual arguments:


The above decision related to something that had no moving parts and required no depth of thought.
Notwithstanding, 42% of the people voted "Yes" for Prop. 91.
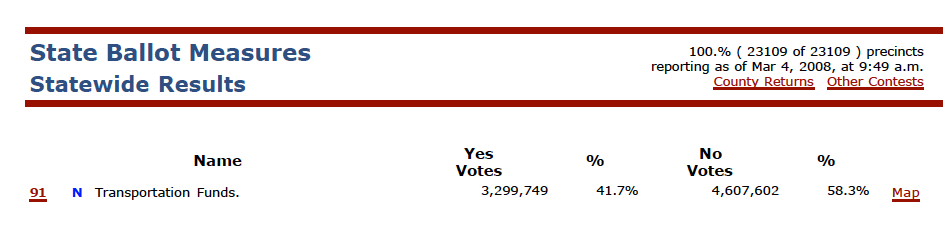
Score one for the "Barely Conscious" category. Also score one for Hollywood, as they may have finally got something right.
If you use the best "standardized thinkers" on earth, but add a few moving parts that require some depth of thought, the situation gets much worse. Glenn Begley of Amgen found that 47 of 53 landmark oncology studies published (89%), primarily from academia, were not reproducible (C. Glenn Begley and Lee M. Ellsi. March 29, 2012, " Raise Standards for Preclinical Cancer Research" , Nature, Vol. 483, 531) . Basically, the research teams proposed studies that were bad ideas (many of which were mechanistically nonsensical in context of known pathways underlying cancer). Those studies were then funded by people that did not understand why they were bad ideas, and when nothing happened, the research teams falsified the data to preserve their reputations and ability to get future funding. The falsified studies were then published by editors of scientific journals that were equally clueless, which effectively diverted future cancer research funding into bad ideas that did not work. Welcome to planet earth.
Let us hope we are never judged by the life form that created the seeds of life. If creating intelligent life in the universe was the objective, the 4 billion year experiment on earth can only be viewed as an abject failure.
The Explosion of Human Life on Earth
Notwithstanding the previous section, humans are what pass for intelligent life on earth.
That minor mutation in glial / astrocyte cell hypertrophy made a major difference in terms of human population hyperproliferation on earth.
Population growth is analogous to cancer growth in that it follows a first order logarithmic progression. The end stages of such a logarithmic progression result in explosive growth in numbers. From the first malignant cell (i.e. a cell that has acquired a mutation in growth control pathways that makes it relentlessly grow and divide), that cancerous cell takes 30 population doublings to reach a 1 billion cell (1 cc) clinically detectable mass, and only 10 more population doublings to achieve a 1 trillion cell (1 liter) mass, which is lethal burden.
It took humans ~ 315,000 years to get through the first 30 population doublings to get to 1 billion people in 1800.
A favorable time in the orbital eccentricity and axial obliquity cycles (Milankovitch orbital warming cycles), use of vaccinations for smallpox (30% childhood mortality), and prosperity from the industrial revolution, all contributed to shorten the next population doubling time.
The next population doubling took 130 years, with 2 billion people in 1930.
A continued favorable time in the Milankovitch global warming cycle combined with the development of nitrogen based fertilizers and mechanization of farming, continued prosperity, the discovery and development of antibiotics, continued development of vaccines ( diphtheria - 1923, tetanus - 1924, whooping cough - 1940, polio-1961, measles - 1963, mumps - 1967, rubella -1969), all resulted in a further reduction in the population doubling time.
The next population doubling only took 44 years with 4 billion people in 1974.
The latest population doubling has taken 46 years, with ~ 8 billion people by the end of 2020.
If you were alive in 1974 and remember the world as being less crowded, it was. There were only half as many people in 1974 as there are today.
The carrying capacity of an environment is defined as the maximum population size of a biological species that can be sustained in that specific environment, given the food, habitat, water, and other resources available.
In 2001 the UN stated the estimates of the carrying capacity of the planet for humans varied widely, with a median estimate of 10 billion, and the top of the range at 16 billion.
We are in our last population doubling, as we will hit the carrying capacity of the earth with this population doubling (if we are to believe the carrying capacity studies).
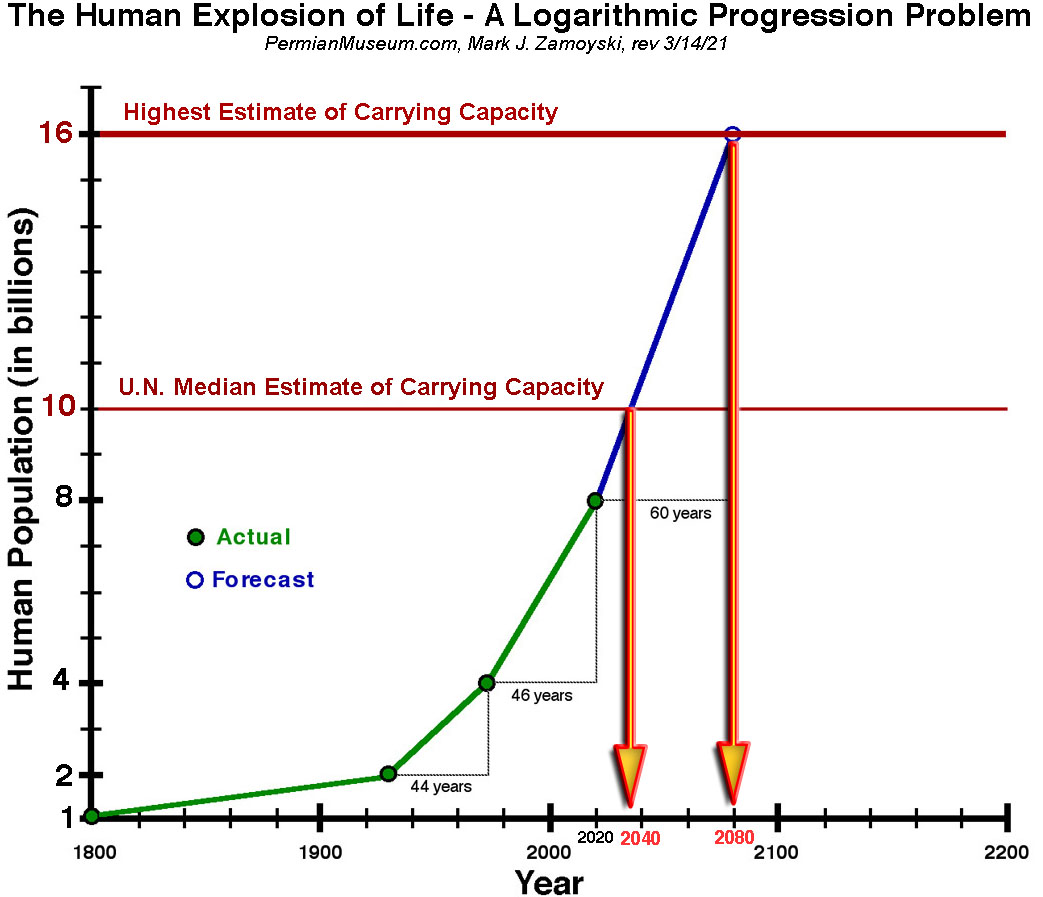
Using our last 2 population doubling times of 45 years, we will hit 10 billion people in around 12 years and 16 billion in ~ 45 years.
The last population doubling saw a decline in annual population growth rate from around 1.6% in the first half of the doubling, to an average of around 1.2% in the last half of the doubling. With 8 billion people now, and a compound population growth rate of 1.2% per year, we would hit 10 billion people in 19 years and 16 billion in 58 years.
Whether we hit the 10 billion mark in 12 or 20 years, or hit the 16 billion mark in 45 or 60 years is not material for our discussion.
The warming part of the earth's orbital eccentricity and axial obliquity cycles is nearing its end and the cooling part of the cycles is next. As temperatures fall, the carrying capacity of the planet falls.
The Milankovitch Orbital Climate Change Cycles and Humans
There is a predictable and sizable human "population reduction" event in our future. It will be caused by the confluence of 1) Humans Reaching the Carrying Capacity of the Planet and 2) The Start of the Cooling Phase of the Milankovitch Orbital Cycle.
Profound Climate Change is routinely caused by the change in earth’s orbit from circular to elliptical (90,000 - 100,000 year cycles) and by variations in earth’s axial tilt (40,000 year cycles), both of which change the amount of incoming solar radiation (insolation), and hence change the temperatures on earth. These warming / cooling cycles are known as the Milankovitch cycles and are explained in the NASA article below:
Milankovitch Orbital Warming and Cooling Cycles

Reference: Full NASA Article on Milankovitch Cycles
While Milankovitch mathematically predicted the temperature affects of these cycles , the Vostok ice core data unequivocally corroborated those predictions. The Vostok ice core drillings for the last 400,000 years reveal the last 4 Milankovitch cycles.
An outstanding Vostok data graphic was posted on the www.brighton73.freeserve.co.uk/ gw/paleo/400000yrfig.htm website, authored by Tom Rees, and a screen shot taken by the author on 10/14/07 is presented below, as the web page could no longer be found.

In summary, the Vostok ice core data showed:
1) The trough to peak increase in temperature during a Milankovitch cycle is ~ 12 degrees C (~ 21 degrees F).
2) We are currently at the peak temperature of the Milankovitch cycle.
3) CO2 levels lag the Milankovitch related temperature changes by about 800 years. That means CO2 increases are the result of the Milankovitch temperature increases.
4) The 3 previous Milankovitch warming cycles increased CO2 levels from ~ 200 ppm to ~ 300 ppm. The "Explosion of Human Life" occurred during this latest Milankovitch peak, and CO2 levels went to ~ 400 ppm in this cycle.
5) What is also known is the CO2-temperature relation. "By itself, a doubling of CO2 in the atmosphere has only a modest effect on global temperatures - an increase of around 1 degree C. Even a quadrupling of CO2 in the atmosphere would only cause a direct temperature increase of about 2 degrees C." ( Lykke Anderson, Director of the Center for Economic and Environmental Modeling and Analysis (CEEMA) at INESAD.) That would mean the additional 100 ppm CO2 would only increase temperature by around one half of a degree C over the previous Milankovitch cycles.
6) What is also known, is this Milankovitch orbital warming cycle will enter its cooling phase next.
Reference: Full Freeserve Article on Vostok Ice Core Data
The graphic below overlays the temperature from the Vostok Ice core data for the prior Milankovitch cycle, on top of the current Milankovitch cycle, to give us a rough idea of what we are in for.
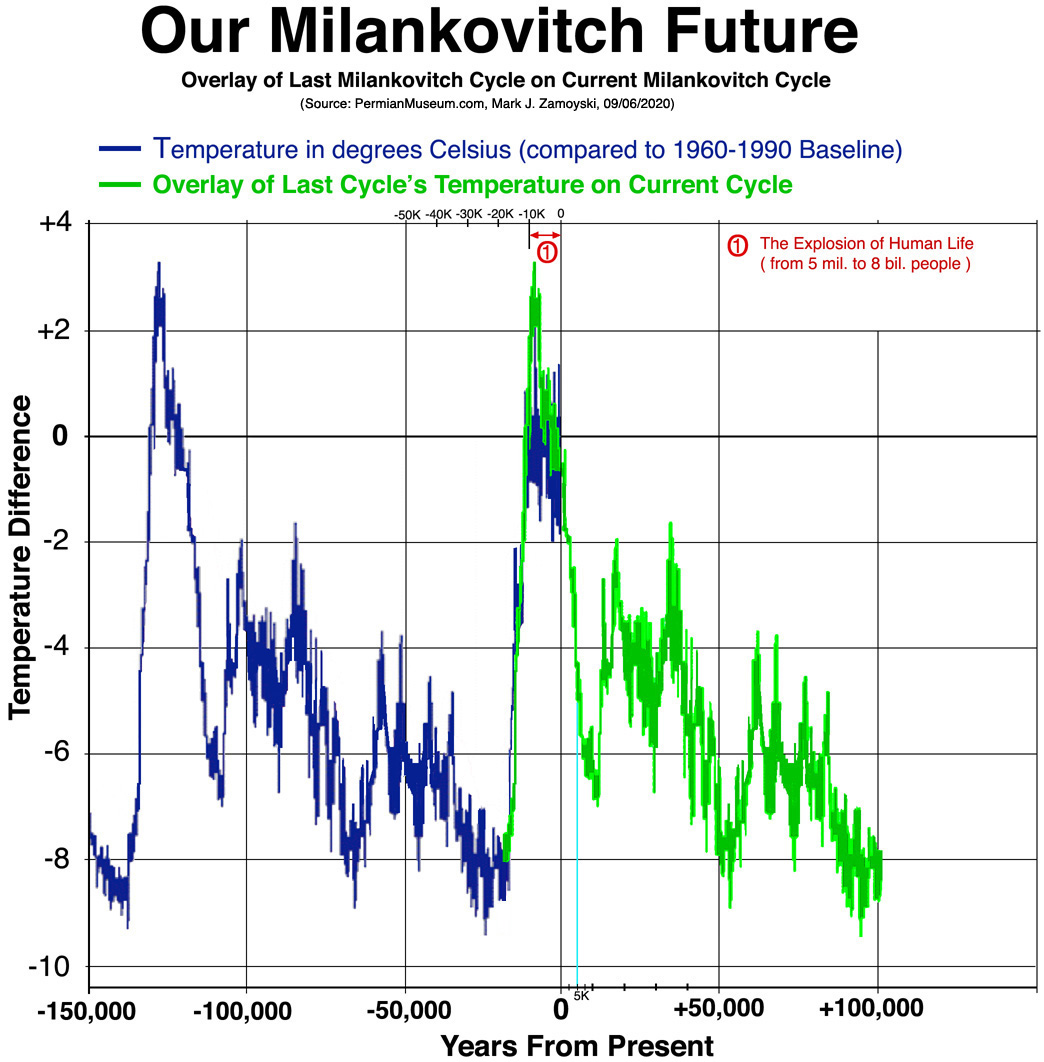
The overlay shows no unusual temperature spikes in the current Milankovitch warming cycle over the previous one. In fact, the current Milankovitch warming cycle did not even reach the peak temperatures observed in the previous Milankovitch cycle.
The overlay also shows the current Milankovitch warming cycle is near its end. We will likely have a few hundred more years of temperatures around the baseline. Then temperatures will fall precipitously, dropping -3 degrees C from baseline by ~ 2,500 years into the future, - 5 degrees by ~ 5,000 years, and -7 degrees by ~10,000 years. Important caveat: this is only a rough estimate, and while the timelines may be off, the magnitude of the drops is consistent with the historical data from the last 4 cycles. More precise timelines would require modeling the 40,000 year axial obliquity cycles over top of the ~ 100,000 year orbital eccentricity cycles.
The Scientifically Nonsensical "Preventing Climate Change" Narrative
The science presented above exposes how scientifically nonsensical statements like "We need to Prevent Climate Change" really are. Let us review a couple of statements often thrown around.
1) RE "Man Made Global Warming": The science presented above shows the rise in CO2 levels from human activity is relatively trivial (i.e. from 300 to 400 ppm is only 100 ppm over historical orbital cycle related increases in CO2 levels ) and its contribution to global warming is equally trivial ( ~ one half of one degree C or ~ 1 degree F). In contrast, the trough to peak global warming caused by solar insolation from the earth's orbital eccentricity and axial obliquity cycles is ~ 12 degrees C or 21 degrees F, and we are currently at the peak as evidenced by the Vostok ice core data.
A synopsis of the Impact of CO2 levels: "An earth with no CO2 would be very cold. The first few tens of PPM produce a strong warming effect, and increases after that are incremental. It is widely agreed that a doubling of CO2 will increase atmospheric temperatures by about 1.2 C, before feedbacks." (Steve Goddard, Anthony Watts, https://wattsupwiththat.com/2009/02/25/a-short-primer-the greenhouse-effect-explained/#more-5853)
2) RE: "We need to Prevent Climate Change": This is a scientifically nonsensical statement on several levels.
Mechanistically, that would mean we would need to "fix" the earth's 100,000 year orbital eccentricity cycles so that the earth's orbit is consistently circular. That would require greatly increasing earth's orbital velocity.
We would also need to somehow "fix" the 40,000 year axial obliquity cycles, or basically straighten the axial tilt, and then prevent the earth from wobbling away from it.
Reducing man made CO2 levels will not change the earth's orbital eccentricity or axial obliquity cycles.
Another huge scientific flaw is that there is no basis for what the "baseline" desired temperature should be. The arbitrary choice of contemporary temperatures is the quicksand the foundation has been built on. Is today's planet, with its frozen poles and cold climate, really better than the Paleocene and Eocene eras that spawned our forerunners, when there was no ice, the planet was covered with forests, global temperatures were 10 - 15 C warmer and CO2 levels routinely ranged from 500 to 800 ppm, with peaks recorded at 4,000 ppm? With earth heading for a cooler climate as humans hit the population capacity of the planet, higher CO2 levels would result in a warmer earth and would actually delay or reduce the size of the human die off that is coming from cooling related reduction of food production and resulting human starvation. That may arguably be considered as "better".
Alternatively, let's take a look at what a warmer world from carbon emissions would look like. Current models predict that a doubling of CO2 levels would only result in a ~ 1 degree C increase in temperature, and a quadrupling would result in a ~ 2 degree C increase. So what would a 1 or 2 degree C rise in temperature mean for you?
While the global baseline (1961 - 1990) temperature was around 16.5 C, temperatures vary widely by geography. This allows you to calculate what a 1, 2, 5, or even 10 degree increase in temperature would mean for you.
Annual Baseline Temperatures for Select Countries
- -16.1 Greenland
- - 5.4 Canada
- - 5.1 Russian Federation
- 1.5 Norway
- 1.8 Iceland
- 2.1 Sweden
- 5.5 Switzerland
- 6 .2 Austria
- 7.0 China
- 7.9 Poland
- 8.3 Ukraine
- 8.5 United Kingdom
- 8.5 Germany
- 8.6 United States
- 9.3 Netherlands
- 9.3 Ireland
- 9.6 Belgium
- 10.7 France
- 11.2 Japan
- 13.3 Spain
- 13.5 Italy
- 15.4 Greece
- 16.4 Lebanon
- 17.1 Morocco
- 17.8 South Africa
- 18.3 Jordan
- 18.5 Cyprus
- 19.2 Israel
- 21.0 Mexico
- 21.3 Bermuda,
- 22.1 Egypt
- 22.7 Hong Kong
- 23.7 India
- 24.6 Congo
- 24.7 Saudi Arabia
- 24.9 Bahamas
- 25.0 Brazil
- 25.3 Puerto Rico
- 25.9 Guam
- 26.3 Thailand
- 27.0 United Arab Emirates
As an example, if you live in the US (8.6 C), UK (8.5 C) or Germany ( 8.5 C), a 2 degree C increase would be like living in France (10.7 C). Five degrees higher would be like living in Italy (13.5 C), and 10 degrees warmer would be like living in Israel (19.2 C). People do live and thrive in Israel, and do not burst into flames when walking outside in the mid day sun.
As another example, if you live in Russia ( - 5.1 C), and are dreaming about a 10 degree C rise in temperature, you are dreaming about living in Switzerland (5.5 C).
However, you may also want to look at the list to see what the temperatures will look like as the Milankovitch cycle enters its cooling phase.
Our Future: Milankovitch Cooling Meets Carrying Capacity
While we have enjoyed the warmth of this Milankovitch cycle, what is inevitably coming is the cooling part of the Milankovitch cycle, as shown in the overlay chart above.
While global warming is life promoting, global cooling is a killer. "A one-degree global change is significant because it takes a vast amount of heat to warm all the oceans, atmosphere, and land by that much. In the past, a one- to two-degree drop was all it took to plunge Earth into the Little Ice Age. A five-degree drop was enough to bury a large part of North America under a towering mass of ice 20,000 years ago." (NASA, earthobservatory.nasa.gov/world-of-change/global temperatures)
The cooling part of the Milankovitch cycle is coming and will drop temperature over the next ~ 5,000 years by around 5 degrees C and by around 7 degrees C in ~ 10,000 years. Accordingly, if you live in the US, UK, or Germany (8.5 C), and wanted to maintain your current "paleoclimate", or possibly avoid living under a towering ice sheet, you would need to live in Italy (13.5 C = +5) or Greece ( 15.4 C = +7). Perhaps the origin of the "cradle of civilization" being in that area is not a coincidence, but a Milankovitch paleoclimate thing.
What is also coming is the human population reaching the carrying capacity of the earth in this population doubling cycle.
Few humans appreciate the explosive numbers inherent in the end stages of a logarithmic progression. Basically, nothing is "sustainable" at our current position in the progression to a terminal condition.
The last two population doublings that took us from 2 billion to 4 billion, then 4 billion to 8 billion averaged 45 years. The last half of the last population doubling saw a slowing down of the annual population growth rate to around 1.2 % per year. At that rate, we would hit the 10 billion median carrying capacity estimate in under 20 years and the 16 billion highest carrying capacity with our latest population doubling, in under 60 years.
Stopping, or reversing, population growth is what is needed to prevent us from exceeding carrying capacity. However, this would likely be opposed by many religious denominations worldwide and politicians opposed to birth control of any kind. It would also likely be unheeded by people that for various reasons need to over procreate.
Increasing carrying capacity by terra formation of planets like Mars is just mathematically too late to be a viable solution. It will only be a short duration fix, at best. Ignoring the lack of a magnetic field, oceans, atmosphere, and the smaller size of Mars, best case, assuming we could transform Mars in 60 years to give it a carrying capacity of 8 billion, in isolation, that would only fix the problem for an additional 30 years. We would be at 16 billion people on earth at that time, and in another half population doubling (30 years) we will have cranked out 8 billion more people and filled up the carrying capacity of Mars.
In 2025 the UN updated its estimate of when we will hit carrying capacity, because of deaths and reduced fertility rates from the COVID mRNA vaccine, that was forced on more than 5 Billion people globally.
The death, disability, and infertility was foreseeable from a molecular biology perspective from inception.
COVID infects only cells possessing ACE 2 receptors, makes them produce a non-self protein, which results in them being killed by your immune system. In contrast, the LNP delivers the mRNA to all 210 cell types, makes them produce a non-self protein, which results in them being killed by your immune system. That killing includes cardiovascular cell, brain cells, nerve cells, reproductive organs, immune system cells themselves, and the list goes on by cell type….
Cancers were foreseeable from carcinogenic DNA templatecontaminants, "badjuvants" like SV40, and use of the fully active spike protein which inhibits DNA damage checkpoints.
Unfortunately, the people that lacked the vision of see the foreseeable, have been appointed to head up the FDA and CDC, and continue to approve mRNA drugs, without having solved the lack of cell specific delivery problem.
The point being, the UN may need to update the 2025 figures again.
When the carrying capacity limit hits, it will begin to manifest as a progressive scarcity of available food. That is the basic math of total available food production divided by the the number of people seeking it. The denominator is growing at a logarithmic rate, while the food production numerator has limited expansion potential, and will actually start to decline as the Milankovitch orbital cooling comes (cold reduces food production).
Food scarcity brings out the most fundamental survival instinct in humans. History shows tribes have killed other tribes over scarce food resources.
History has also shown that governments use food scarcity as a weapon against its citizens. Stalin's Holodomor (terror famine) during the food shortages of 1932-1933 was used to starve an estimated 5 million dissidents by withholding food distribution to Ukraine. After Roosevelt and Churchill gave Stalin several eastern countries in 1945 at Yalta, Stalin used the starvation and slavery of gulags (political labor camps), in addition to direct executions, to murder an estimated 20 million dissidents in those countries. Welcome to planet earth's ruling elite.
Hopefully, the above has given you an appreciation of the magnitude of the problem that is coming.
Well, that pretty much brings us up to date on 4 billion years of life on earth, as well as a neighborhood crowding forecast for the next hundred years and a weather forecast for the next 100,000 years.
Chapter 8
Interstellar Visitor Video Gallery
In 2024, a soft X-Ray mapping of the visible universe by the eROSITA satellite discovered an "interstellar tunnel network".
In August 2025, Rep. Anna Paulina Luna disclosed that the government had known of the existence of inter-dimensional beings that had visited earth.
Infrared security footage at our site in 2017 picked up a series of events that may now be interpreted as arrivals from an off-ramp of the interstellar highway.
Interpretations are purely those of the Author, and may or may not be correct. This chapter is intended to provide that footage for other scientists to interpret and draw their own conclusions.
Form Author's perspective, the story comes in three parts:
1) The Scout (1-19-2017)
2) The Scouting Party (5-19-2017)
3) The Plasma Portal (summer of 2017)
Below is Part 1. Parts 2 and 3 are under construction.
Part 1: The Scout
One camera picked up the plasma trail of something coming in fast and hot (upper left corner of frame).
A frame by frame reveals it passed through in a single frame, and the residual heat trail dissipated quickly over the next few frames.
Frame extracts from "Incoming":

Please keep in mind, the infrared shows the effect of the entity on the ambient environment, and not the entity itself. This is analogous to the wake of an invisible boat that passed through the water - the wake does not show what the boat looked like, just that it came through
The actual clip: Incoming Video

Another camera was just above the impact site, and picked up the Post Impact Transformation and more deliberate subsequent movement of what came through.
Frame extracts from "Post Impact Transformation":

The video clip itself: Post Impact Video
Part 2: The Scouting Party
UNDER CONSTRUCTIION
Part 3: The Plasma Portal
UNDER CONSTRUCTIION
Chapter 9
Galactic DNA Transport
Added April 2026, © 2026, Mark J. Zamoyski
Introduction and Background
Earlier chapters focused on the life forms spawned by the bacterial DNA that arrived from the asteroid impacts on earth 3.8 Ba and looked at the resulting life forms it spawned.
This chapter looks at fossilized life forms from a piece of another planet for comparison.
A carbonaceous chondrite meteorite was found in the Permian soil about 20 miles away from our earth fossils site.
That means a chunk of a foreign planet arrived in our solar system 4.5 Ba, where it became part of an asteroid, a part of which then arrived on earth around between 250 - 300 Ma as a carbonaceous chondrite meteorite.
From a life science perspective, that means it brought prokaryotic DNA from a foreign planet, as well as a fossil record to show the life forms spawned by that DNA. It is the ultimate time capsule from another planet, or at least from a small part of that planet.
This allows us to compare life forms spawned by DNA on other planets, versus those spawned by the DNA that arrived on earth 3.8 Ba.
Let's start by summarizing what we saw happen on earth.
A BRIEF HISTORY of DNA on Earth:
Three explosions of life occurred from the prokaryotic DNA that arrived on earth 3.8 Ba:
1) The Prokaryotic (Bacterial) Explosion of Life: The prokaryotic DNA brought mitochondrial DNA to earth 3.8 Ba. Mitochondria is a cell’s power plant that stores energy from aerobic respiration (metabolism of glucose). Human mitochondria is bacterial - circular DNA, separate from the nucleus, and its nucleotide sequence points back to a DNA reassortment of rickettsia, rhizobacteria, and agrobacteria.
Physiological life can be defined as a collection of concentration gradients. Their presence means life, their absence means death. It takes energy to maintain concentration gradients. Without the mitochondrial DNA that arrived on earth some 3.8 Ba, life as we know it today would not exist, including humans.
Bacteria was the most successful explosion of life in earth's history. On planet earth, bacterial biomass exceeds that of all plants and animals combined. One gram of soil contains 100 million to 1 billion bacterial cells.
2) The Unicellular Eukaryotic Explosion of Life: Eukaryotic cells appeared 1.5 Ba and contain only a subset of prokaryotic DNA.
Eukaryotic cells only had a single self-assembling lipid bilayer cell wall membrane, rather than the though triple layer lipid - protein - lipid prokaryotic cells.
Eukaryotic cells had fragile multiple strands of DNA rather than the structurally stronger circular DNA. DNA efficiency was also much worse, as measured by the number of proteins they could produce per million base pairs of DNA.
However, the single self-assembling lipid bilayer cell wall membrane and more fragile DNA both facilitated DNA reassortment of eukaryotic cells into new life forms, under the right conditions.
The most significant eukaryotic lineage spawned was the Calcium Secreting Filter Feeders (CSFFs) that lived in colonies and built calcium structures that were able to host DNA reassortment in shallow coastal oceans (Ch. 3). CSFFs also brought early bone DNA.
3) The Multicellular Eukaryotic Explosion of Life - 500 Ma (Cambrian) and 250 Ma (Permian):
The earliest known multicellular life forms were microscopic nematodes (roundworms and eelworms) that appeared more than 500 Ma. They are basically a mobile intestine, with a mouth, anus, and gonads (cells that produce haploid cells). They had a protective protein skin and two neurons running the length of the body connecting to muscle that allowed fish like motion. The mobile intestine with gonads went on to become the core blueprint for all multicellular life forms that followed, including humans.
Nematodes provided digestive, reproductive, muscle, nerve, and skin DNA that was now readily available for the DNA reassortment process that created new life forms.
The Cambrian Explosion of Life had unicellular and nematode DNA to work with to generate new life forms by DNA reassortment, followed by 250 million years of evolution.
The Permian Explosion of Life had the same input, plus the evolved Cambrian life forms that survived the Permian extinction events. The survivors included sharks, sting rays, reptiles and lung fish. This allowed a more robust explosion of life that gave rise to the age of the dinosaurs and winged flight.
Life Becomes Aware:
The appearance of a brain cavity that was larger than necessary to house just neurons, combined with either a DNA reassortment or birth mutation, spawned the hyper-proliferation of astrocyte cells that interconnect neurons in a way that allows for propagation of the calcium ion wave which hosts the measurable electromagnetic phenomenon underlying consciousness and memory formation.
Neurons make up 97% of a Leech brain. In contrast, neurons only make up 10% of a human brain, and glial / astrocyte cells make up the rest. It made a big difference.
However, addition of a brain to a mobile intestine with gonads would foreseeably result in a conflicted road toward the development of intellect.
DNA Transport:
Carbonaceous Chondrites are the Galactic DNA delivery vans.
Earth is 100% extraterrestrial as it cannot make elements. It only re-arranges elements received.
Elements are made by stars and supernova explosions eject these elements through space at 10% the speed of light (30,000 kilometers per second).
Supernova explosions also directionally emit lethal amounts of gamma and x-ray electromagnetic radiation and the high speed ejection of matter creates a shock wave that propagates throughout the interstellar medium.
Any planets in the cross hairs of the directional gamma and x-ray emission would likely be sanitized of surface life.
Any planets clobbered by a glob of Supernova ejecta moving at 10% the speed of light can it turn have chunks the planet itself ejected into space at high velocity and become part of the debris stream that is eventually accreted into asteroids, and chunks of which fall to earth as meteorites are called Carbonaceous Chondrites.
Packaging and Transport of DNA:
While both prokaryotic (bacteria) and eukaryotic DNA from planet ejecta would be transported, the reasonable expectation is that only the prokaryotic DNA would arrive in a viable form. Prokaryotic DNA is what spawns the highly "DNA devolved" eukaryotic life forms that inhabit a planet.
Prokaryotic cells have a tough triple layer cell wall, can thrive near volcanic vents 3,500 feet below the ocean surface and live two miles deep in soil at pressures of 5,000 PSI. In contrast, eukaryotic cells have a single, fragile lipid cell membrane that can be easily ruptured by a soft punch.
The sheer numbers of prokaryotic cells virtually eliminates the possibility of total eradication. On planet earth, bacterial biomass exceeds that of all plants and animals combined. One gram of soil contains 100 million to 1 billion bacterial cells. Coastal oceans contain 1,000,000 cells per ml.
Prokaryotic cells also have highly stable and efficient circular DNA, that cranks out ~1,000 proteins per million base pairs of DNA, versus human eukaryotic DNA strand junkyard and only crank out 7 proteins per million base pairs, and 98% of which in non-coding (not used). It makes sense to transport the "DNA gem" and not the "DNA junkyard".
Most important, prokaryotic DNA has all of the sequences for spawning unimaginably biodiverse new eukaryotic life forms, including the DNA for the eukaryotic cells that do that.
Other than the tough prokaryotic cell wall, the next level of packaging of prokaryotic DNA for transport is the meteorite itself. While a meteorite enters earth's atmosphere at 12 to 40 kilometers per second (~ 27,000 to 90,000 MPH), the impact speed is generally between 200 - 400 MPH (322 - 644 KPH) because of the atmosphere.
The surface temperature of the meteorite can reach 3,000 F (1,650 C) from the ram pressure during flight through the atmosphere, however that is limited to the fusion crust, and the core itself remains at a temperature similar to that of deep space (very cold). Meteorites that reach the ground are typically cold.
Additionally, some of the extraterrestrial fossils we will review shows a third level of "survivability" packaging for bacteria - the internal gastrointestinal "cushioning" provided from "multicellular life form packaging".
Photo Gallery of Meteorite Fossils
All photos in Ch. 9 below - Copyright 2026, Mark J. Zamoyski
The carbonaceous chondrite itself is a raggedy looking rock after 250 - 300 million years of weathering on earth. However the sparkles, or shiny flecks, gave it away on a sunny day. We have named it "Life Rock".

It should be noted we are doing only a surface examination of the carbonaceous chondrite, with a simple Bebang 1200X digital microscope (~ $150 through Walmart).
Some life forms are morphologically similar to their "super sized" versions on earth. The morphological form speaks to the underlying DNA. The tiny size is not yet understood, but may speak to life on a high gravity planet or some different growth control pathways than our own.
The preservation in life forms deeper in the rock is conventional. The preservation of life forms at the surface is lifelike, and may speak to a supernova gamma ray burst.
The DNA underlying the fossils on the meteorite fills in a lot of missing pieces in our normal size earth fossils found at the CSFF site 20 miles away. Together, the two provide a more complete picture of life forms that exist on earth today.
Your first step onto the rock at ~ 1000 X magnification reveals an unimaginable world:

You will see shiny metal fragments, calcium-aluminum inclusions (CAI) , and the 3 main food sources on that part of planet (red "fruit" pods, brown "ranch dressing" pods, and dark shelled clams).
The shiny metallic flecks you see on a sunny day are primarily nickel-iron alloy, (mostly iron with 5% - 30% nickel) called kamacite and taenite. The metal comes from the supernova explosion and the metal flecks condensed 4.5 Ba directly from the hot gas of the solar nebula. Chondrites have 2 - 15% metallic content by weight.
The Calcium-Aluminum-rich Inclusion (CAI) is typically found in carbonaceous chondrites and was formed by condensing hot gas in our protoplanetary disk 4.5 Ba, per current reference sources.
The ground you are walking on is 45% Olivine, 25% Pyroxene and 10% Plagioclase, minerals capable of forming crystals under the right conditions, so expect to see some nice crystals lying around, particularly after 300 million years on earth that included a stint in the ocean and tectonic activity that lifted the rock to 4,000 feet above sea level.
A good place to find life forms is where the food is, so let's start our walk about by looking at what is eating the red "fruit" pods.
The first red fruit pod we see is being eaten by a beaked spider with a spherical body:

On closer inspection, it appears the spider has 4 legs on its side, and 2 larger legs on its back that allow it to brace itself on the fruit casing.
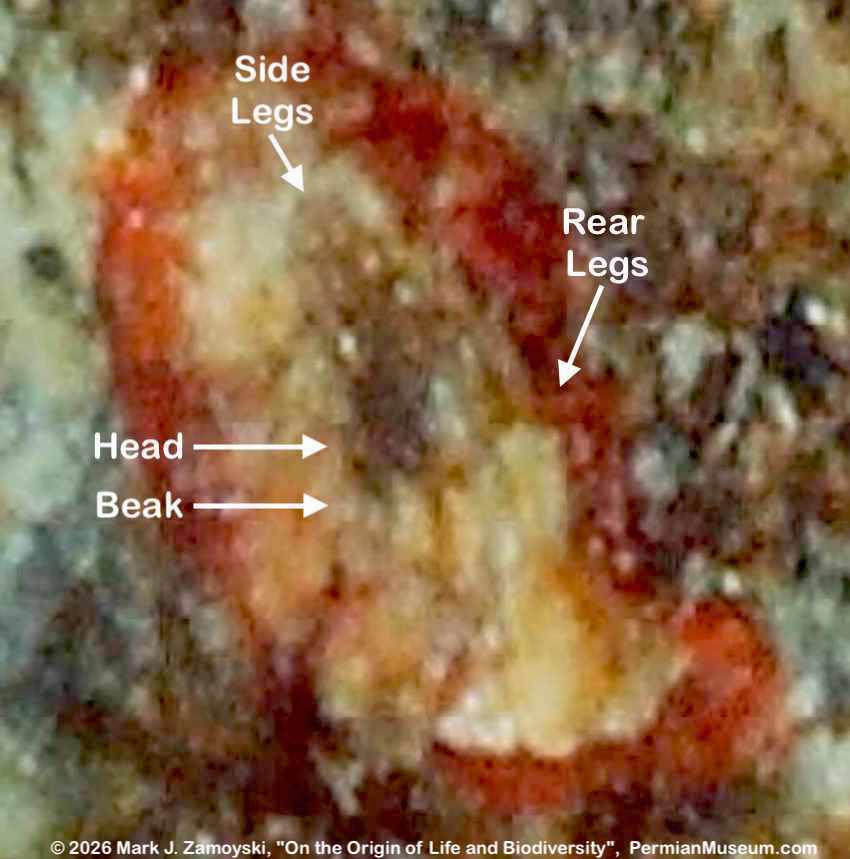
The next fruit pod appears to contain a cockroach shaped insect below, and a spherical in now on top of it, likely eating it instead of the fruit. The beak and head of the spider are obscured by a vertical anatomical protrusion from the submerged insects back.
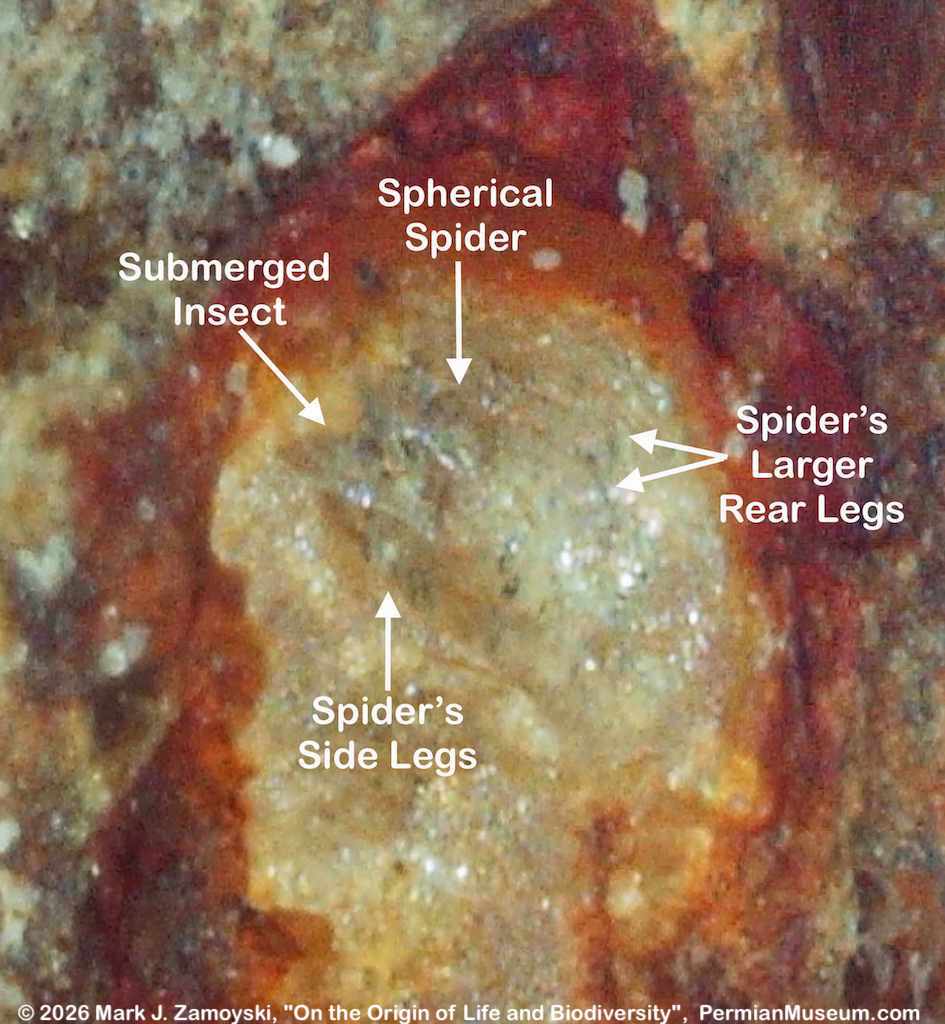
The spider appears to be have its 4 side legs wrapped around the submerged insect, to hold it in place while it eats it, and the spider's larger rear legs appear to be for pulling it out of the fruit pod when it is finished. The total leg count of 10 is in contrast to 8 for an earth spider.
The next fruit pod contains a life form not known to us:
In the fourth pod we see a greenish frog or salamander-like life form that is using its right arm to brace itself on the fruit pod wall, while using its left arm to reach down and retrieve a submerged life form:
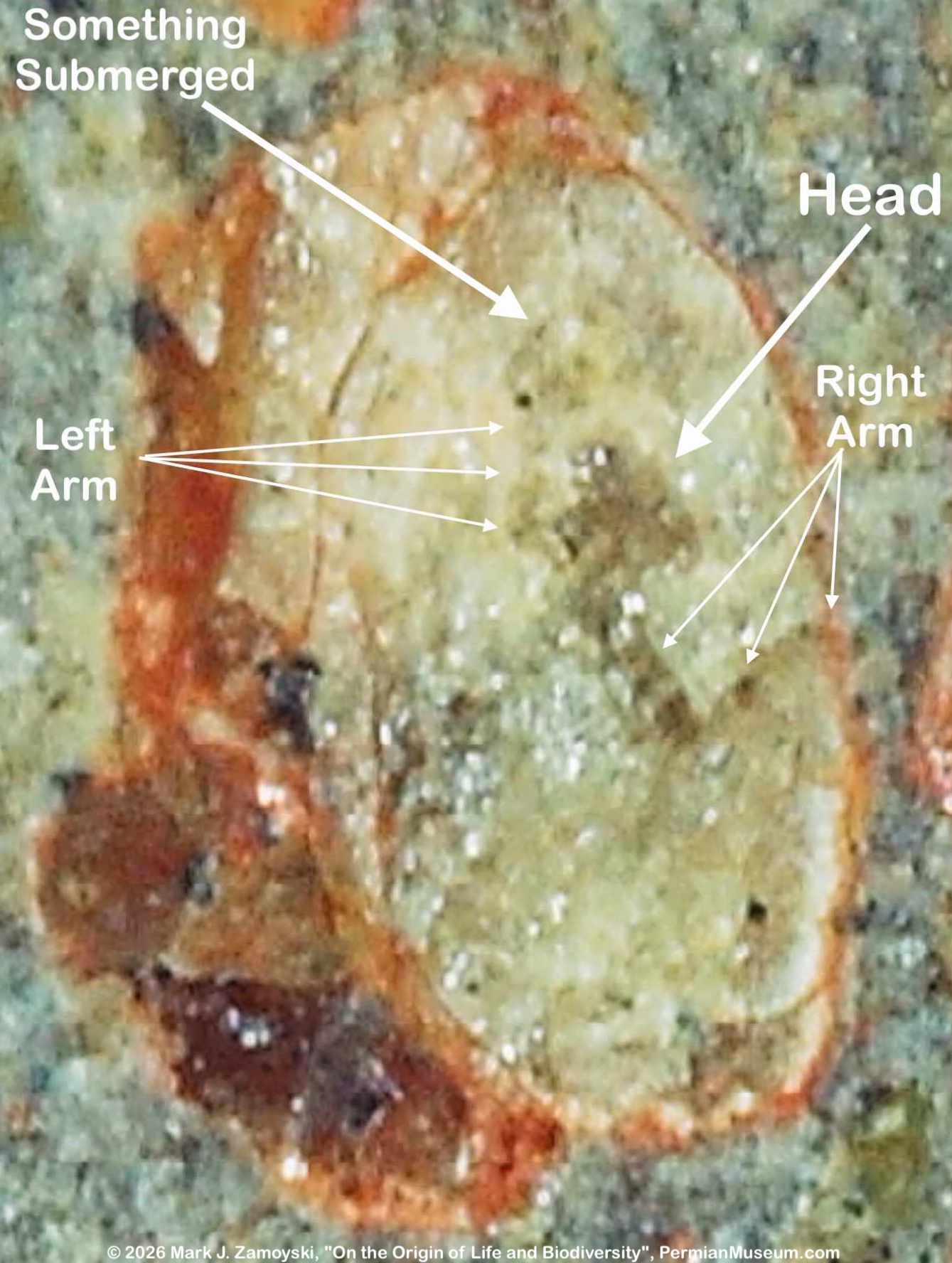
Now we are going to stroll over the the brown pods to see what they have inside, and what is eating them.
We find an open brown pod and see that what is inside looks like Ranch Dressing:

There is an abundance of life forms we will call "Red Eaters, Metal Accumulators" or REMA's, that feed on the Brown Pod contents.
There are variations in their mouth structures, and they may have either two or three compartments / segments, but appear to share the common feature that they appear to accumulate metallic content in their compartments.
A photo of one type of REMA, with the background excluded is shown below:

Adding the background back in shows the REMA, mouth open, making a dash for a Brown Pod:

The photo of another type or REMA shows its face buried in the brown pods or possibly the dark clams. It actually appears it may be eating something that was eating the brown pods or clams.

The below photo shows what appears to be a baby REMA that has no metallic accumulations.
The next photo shows a maturing REMA that appears to have metallic accumulations starting in the front most compartment, but not yet in the rearmost compartment:

What appears to be a more mature REMA is packed with metallic material and looks more like an aluminum recycling truck than an organic life form:

The Red Eaters also include a type that appears to eat other Red Eaters. The Red Eater Eater appears on top in the photo below, appears to have a squid like beak, and appears to be eating the red eater below it. The red eater below appears to already have had some bites taken out near its rear.

In context of Red Eater Eaters, the metallic accumulation between the internal organic structures and the exoskeleton now makes sense. It may form a "armor" barrier against the Red Eater Eaters, or at least a un-tasty mess to go through to get to the organic part. There does appear to be preferential skin or exoskeletal eating going on in some of the photos.
Leaving the brown pods, we can see some of the numerous other life forms on the planet:

Just like on earth, life forms try to protect themselves from predators by seeking shelter, such as the life form in the crevice just to the left of the CAI.
Taking a moment to enjoy the landscape, the metal / gem combinations on chondrites can be quite spectacular, as shown in the photo below.

Shrimp-like life forms also inhabited the planet:
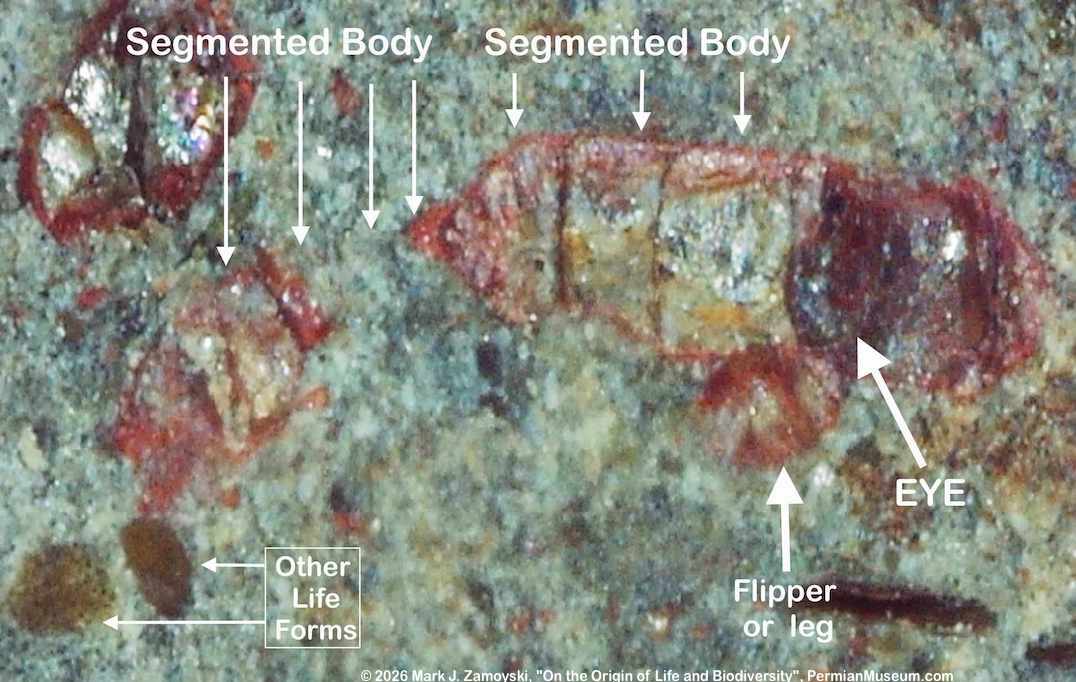
Reptiles also appear on the planet. The first photo is what looks like a partially submerged crocodile:

The fossilized Lizard below show an exposed rib cage and GI tissue. This shows the feasibility of prokaryotic DNA transport in the intestinal tract of life forms, and not just in the soil.

The last two photos are of a mammal life forms on the planet.
The first is a rodent resembling a rat eating a primitive life form that resembles a orthocone or leach.

The last photo shows a more eroded part of the rock where a fossil of that rodent appears to be emerging from deeper (presumably older) part of the rock:
Both photos show the three digit hand comprised of two fingers and an opposable thumb, and not the 5 digits typically found on earth.
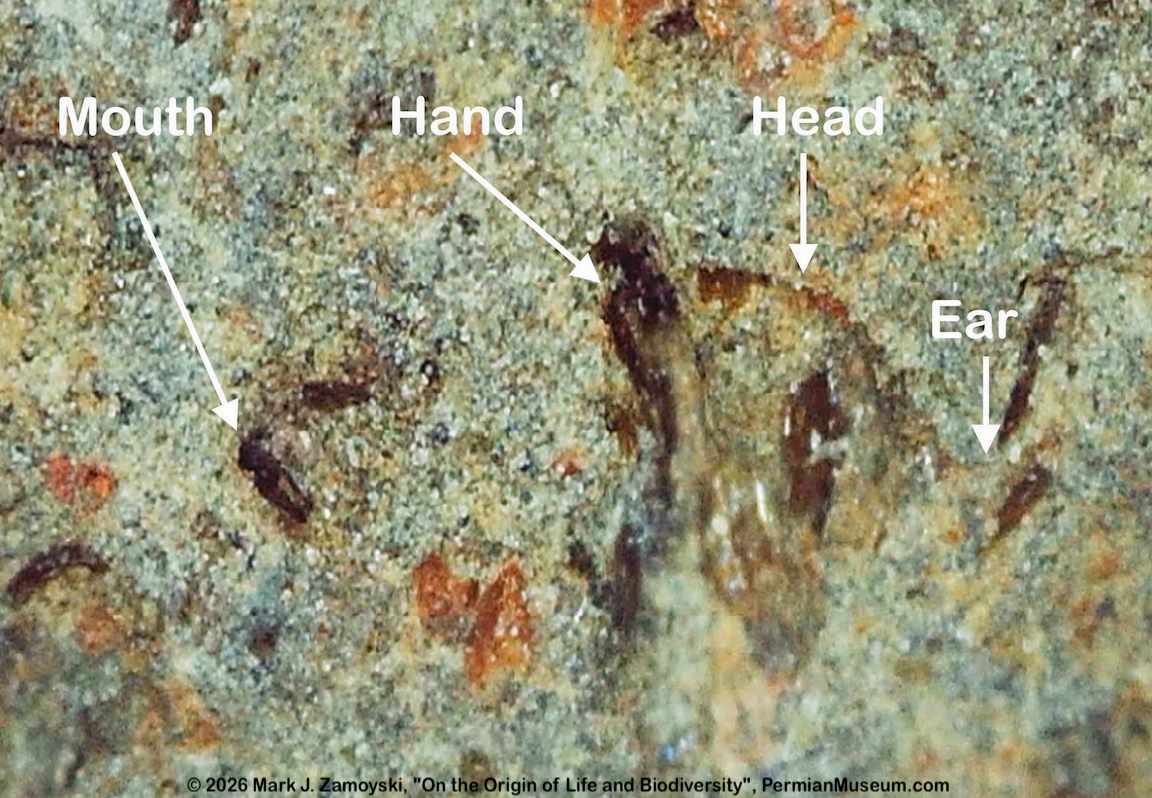
Observations / Conclusions
Important Background: The earth fossils presented in our book and the meteorite fossils presented in this chapter were both found in Permian Soils (~ 250 - 300 million years old), however the two sites where ~ 20 miles apart.
The important differences between the life forms found in earth fossils versus the meteorite fossil:
0) The first eukaryotic multicellular life forms found on earth were microscopic nematodes, which served as the core blueprint for all multicellular life forms that followed, all the way through to humans. We found no nematodes on the meteorite, but since the scaling of life forms on the meteorite is 1000X smaller, nematodes would be sub microscopic so we would not see them.
1) We never found a spider fossil at the CSFF rich earth site. The meteorite had two spiders.
2) We never found mollusks. The meteorite had an abundance of mollusks (the dark clams).
3) We never found an amphibian like a frog. The meteorite had a frog in action.
4) We found an abundance of avian forerunners in the CSFF fields. The meteorite had no avian forerunners.
5) The reptilian fossils we found at the CSFF rich site were forerunners of reptilian carnivorous dinosaurs. The meteorite reptilian forerunners were conventional lizards and crocodiles.
6) The mammal fossils found at the CSFF rich earth site were forerunners of certain dinosaurs and early primates. The meteorite's mammal forerunner was a mouse or rat like rodent.
In context of what was covered previously, the meteorite would have the prokaryotic DNA that spawned the eukaryotic life forms seen on that meteorite.
It also suggests DNA reassortment events may have varied at each active CSFF site. Our earth fossil site had sting ray and jelly fish DNA, and saw life forms like fish and aquatic birds with contractile propulsion sacks like a jelly fish. Water wings (sting ray) were basically a forerunner of air wings.
As the meteorite DNA would have found its way into a nearby CSFF field, it would get shredded and reassorted, recreating the same or similar cells to those it previously generated, plus variants as it combined with the indigenous DNA.
That would have added rats, spiders, frogs, crocodiles, etc… that we did not see at the earth fossil site.
Basically, the ability of meteorites to bring new DNA to a planet, and have that DNA be integrated into a planet's playbook of life, is mechanistically possible, but dependent on that DNA making its way to an active CSFF colony, or functionally equivalent DNA reassortment cavitation structure, moving water through those structures, and an extinction event that allows the new life forms to survive and thrive in the absence of established predators (Ch. 3).
This meteorite arrived on earth at exactly the right time and at the right place in that respect.
1) There were extensive CSFF colonies in action, as evidenced by the abundant CSFF fossilized structure found and presented in the book.
2) The part of Arizona where the meteorite fell was a shallow ocean at the time, which provides the movement of water containing the prokaryotic cells (1 mil/ml), allowing CSFF structures to shred them and create DNA reassorted eukaryotic cells. The unused shredded material becomes a nutrient rich environment (like an egg) that allows the reassorted cells to thrive.
Based on data published by Ron Blakey and Wayne Ranney in "Ancient Landscapes of the Colorado Plateau", Arizona had beachfront property.
Arizona Landscape
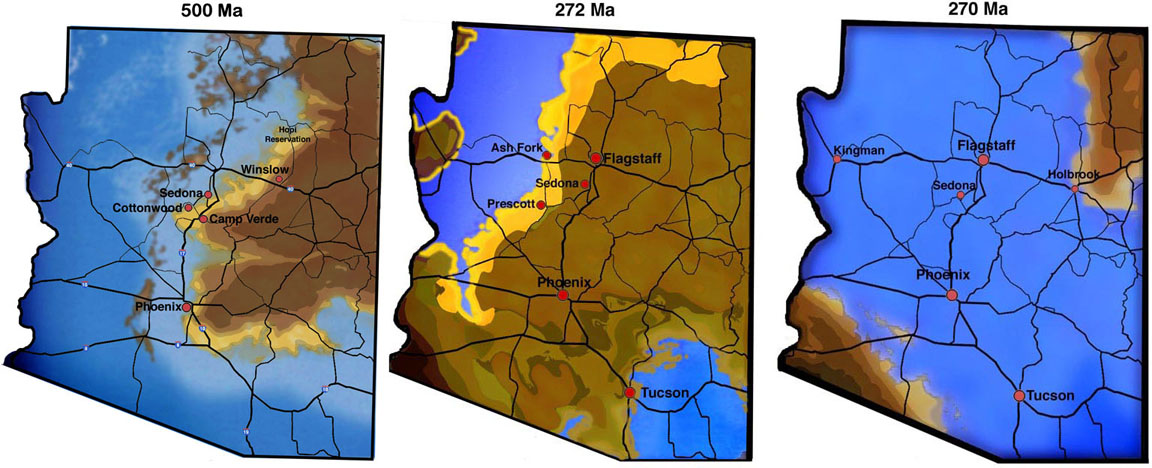
3) There were 4 extinction events during the Permian period, with the last and most severe killing 90% of ocean species. Extinctions are a requisite condition for survival of new life forms, otherwise they are just lunch for an established predator.
The below graphic depicts these pulses for terrestrial tetrapods, based on data published by Sahney and Benton (Sahney, S and Benton M. J., “Recovery from the most profound mass extinction of all time”, Proceedings of the Royal Society B, 275, 759 - 765 , 2008)
Permian Extinction / Recovery Pulses
Millions of Years Ago (Ma)
Extinctions plus CSFFs plus shallow oceans allow for another "roll of the dice", but this time with both pre-existing DNA, and new DNA brought in by meteorites.
The Permian period had 4 rolls of the dice, culminating in a new cast of characters, with dinosaurs becoming the dominant life forms on the planet. Dinosaurs would dominate the earth for ~ 180 million years, until an entire asteroid fell to earth 66 Ma.
The global cooling that followed wiped out terrestrial life forms over 25 kilos (55 pounds). That left the smaller mammals (including primates), lizards, and birds to inherit the earth.
The primates won.
Photo of earth primate (250 Ma) found in the Permian CSFF fields:
Evolution cannot create new life forms. It only makes small changes to an antecedent lineage from selective advantage. Birth mutations and cross breeding can result in big leaps in a lineage.
By 17 Ma the primate lineage had chimpanzees, which have 98% the same DNA as humans. The Homo ("man" in Latin) genus first appeared ~ 2 Ma with Homo Erectus, which went on to spawn the Homo Neanderthalensis and Homo Sapiens (anatomically modern humans) 300,000 years ago. An astrocyte hypertrophy birth mutation changed everything. Astrocyte cells are responsible for propagation of the calcium wave in the brain, which in turn hosts the measurable electromagnetic phenomenon underlying consciousness and memory formation (Ch 5). Humans went on to dominate the planet.
The DNA of mammals and reptiles is 89% the same. Had the series of favorable birth mutations happened in lizards rather than primates, the lizard people would be the dominant life form on earth.
As a final point, the fossilized life forms brought by the 4.5 billion year old meteorite, while different, are at about the same stage of DNA Reassortment and Evolutionary development as our Permian earth fossils. The point is that the other planet had a 4 billion year head start on us, and only had two choices: lizard people or rodent people. Hopefully the Lizard's won.
Important Disclosure: We do not have access to the expensive equipment available to academia, and our observations and conclusions are based only on simpler methods available to us (USGS surface soil age maps, specific gravity measurements, magnetic attraction testing, and both visual and microscopic examination of the meteorite surface for the presence of metallic fragments, CAI's, and fossilized life forms).
The rock was on earth for 250 - 300 million years, so it is possible that indigenous microscopic earth life forms, 1000 X smaller, but morphologically identical to their larger counterparts on earth, inhabited the surface of the rock, and at some point were immaculately preserved by a instantaneous fossilization event. However, the probability is extremely small. Add to that the ecosystem itself seen on the rock, which includes 10 legged beaked spiders, 3 fingered rodents, treeless fruit that grows directly from the ground, and life forms that eat and accumulate metal, etc …, and the probability that the life forma are terrestrial is infinitesimal.
Photo Gallery of Sectioned Carbonaceous Chondrites
Background
Life Rock came to earth 250 - 300 Ma with 2 traveling companions, also carbonaceous chondrites we will call "Big Bob" and "Freckles".
All 3 landed in the ocean next to each other. They watched tectonic activity lift the ocean floor, and found themselves in a river as the water drained away and lifted them to 4,000 feet above sea level.
Carbonaceous chondrites contain organic material. Their specific gravity ranges from 2.5 - 3.0 grams per cubic centimeter.
Ordinary chondrites are not supposed to have organic matter, but mostly accreted "star stuff" from supernova explosions and have a specific gravity of 3.0 - 3.7.
Planetary crust material is lighter because planets are molten when they form, and heavier element like iron and nickel sink to the core. Limestone (calcium and carbon based) ranges from 2.5 -2.7 and Sandstone (silicon) ranges from 2.6 - 2.95. Chondrites contain 2 - 15 % iron/nickel alloys, which are in the 8 - 9 specific gravity range.
Life Rock was a lightweight with a specific gravity of 2.5, closer to typical planetary crust material's specific gravity. "Big Bob" is a heavyweight at 3.0.

Sectioning of Big Bob reveals both life forms, as well a really nice metallic inclusions, which will be shown in the photo gallery.
"Freckles" is more interesting as the 'freckles" appear to be millions of years of erosion washing away matrix, exposing inclusions and chondrules the were once inside. This is particularly important, as if inclusion prokaryotic DNA is now released into the native soil, it becomes part of earth's future playbook of life.
Accordingly, we sectioned Freckles heavily, to see what DNA it released on earth.

Freckles contains some of the same life forms as Life Rock, but also additional ones, as will be shown in the photo gallery.
Caveats
Life forms seen in sectioned slices are nowhere near the clarity of the surface dwellers seen on Life Rock.
1) Dead Bodies: A live body in action is easily recognizable. Dead bodies, randomly positioned, are not as recognizable.
2)"Luck of the Cut": Sectioning provides a view in a single plane. If a life form is oriented perpendicular to the saw blade, you won't likely understand what you are seeing.
3) Orientation within the plane: Bodies in space have no up or down or font to back when they get accreted into an asteroid. A 360 degree rotation within the plane is required both to identify them and then to photograph them in the most understandable position.
Because this is an enormously time consuming process, we have started with a fast drive-by on one side of one slice from Big Bob and one side of one slice from Freckles. This is the stories they have to tell so far, starting with Big Bob:
The Sectioned Carbonaceous Chondrite Photos (~ 1000X mag.)
The first photo shows a carbonaceous chondrite can include what looks like a piece of an ocean floor that dried out, with fragments of what lived there.

The next photo shows it can also include skeletal remains.

It can also contain reptilian or amphibian remains that include soft tissue.

Carbonaceous chondrites can also include planetary ejecta like fully preserved subterranean life forms, right along with the metallic ejecta right next to it.

The life form above appears to be like one that we saw being eaten by a Red Eater on Life Rock. It would make sense, as all 3 meteorites arrived together, and likely from the same place the planetary ejecta found on Life Rock came from.
The brown pods with ranch dressing from Life Rock can also be found as inclusions in the other carbonaceous chondrites. This expands the possibility of DNA transfer to include plant life.

Eggs and virtually any other types of life form DNA from planetary ejecta seems to be on the list of possibilities.

The first photo in the Freckles slice is a "luck of the cut" trifecta.
There were two segmented "eaters" that were not quite perpendicular, but not quite parallel, relative to the saw blade, so we actually dissected the tail end of the top one diagonally through the life form, and trimmed off the exoskeleton on the tail of the lower one. That revealed a wealth of anatomical information.
The 3 things it shows:
1) The red eaters on life rock were actually brown, so the red color was likely part of the manner of death, just as we saw with the rat like rodent. On possibility could have been a direct hit by the supernova Gamma/X burst on Life Rock.
2) The segmented structure in these life forms is actually like the fuselage of a plane - structurally strong rings, with a skin exoskeleton attached to them.
3) There is an unobstructed lumen that runs through the center of the life form. It appears to go into a structure with many folds, resulting in a large surface area, presumably for extraction of oxygen from water. We have called it "Gill- Lung", as it is internal and singular (no bilateral symmetry).
The photo as we saw it:

The photo as we currently interpret it:

It takes a while to get used to looking at things in a single plane, and trying to fill in the unseen parts normally seen in a 3 dimensional world and from an understandable viewing perspective.
As an example below, the photo may be confusing at first glance.
The annotated photo below shows how once you orient yourself on a few known features, you can fill in the rest that is outside the visible plane.

In this example, if you realize you are looking from the top at a dinosaur like life form, you can expect the legs will be in the matrix and not be visible, and that because it is diagonally oriented the part closest to you may be clipped by the saw blade, either exposing or removing part of the life form. Fortunately, in this case. it just sliced off the skin and tissue on the lowermost part of the tail.
We will provide both the raw photo and our interpretation of the photo in cases like this. You can decide for yourself. Our interpretation may not always be the correct one. A single plane view may have multiple interpretations, particularly when a life form is perpendicularly or diagonally sectioned or from a bad viewing perspective.
While life forms like reptiles may not seem significant, keep in mind, the DNA of mammals and reptiles in 89% the same.
The most abundant planetary ejecta on Freckles are amber pods that are filled with white ranch dressing like ooze, just like what we saw in the brown pods on Life Rock.
Many of the amber pods are broken open, as shown in the photo below. It is not clear if that was from the pod's impact during the accretion formation of the asteroid, or by a subsequent impact by a metallic fragment (from the supernova ejecta) being accreted onto the asteroid.
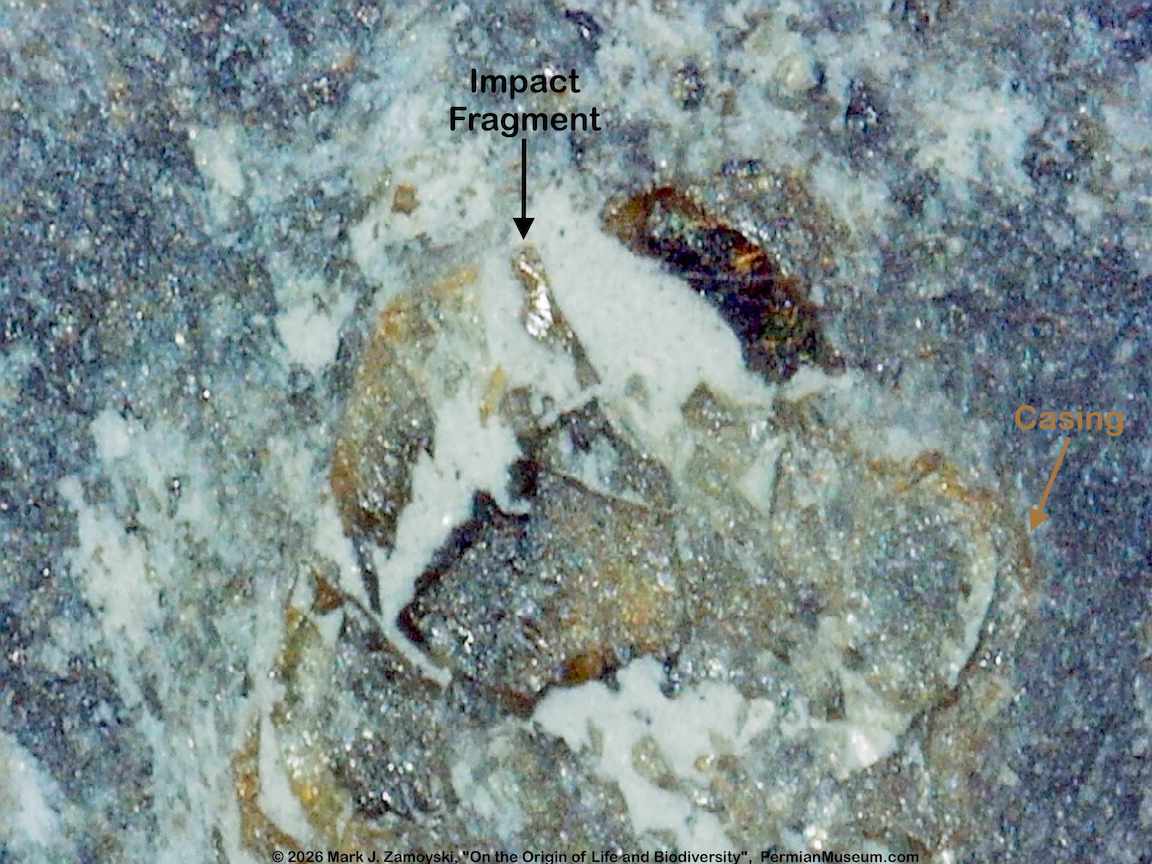
The photo above is typical, a broken casing, with a metallic fragment that was likely a perpendicular hit. The specks in ranch dressing appear they could be the centers of seeds.
There are also some pods that appear to have been sliced through by a metal fragment. This type of impact appears to leave a smear or "Drag Track" of the white substance, with the impacting object at the end of the Drag Track. There appear to be two parallel tracks in the photo below.

The incoming fragment on the left appears to have hit the darker pod/clam area first, then went through an amber Pod 1 with minimal damage, before coming to a stop in the upper right corner. The second incoming appears to have hit and obliterated amber Pod 2.
A zoom-in reveals the reason why.

The incoming fragment on the left hit the root of Pod 1 and went straight through. The roots apparently do no contain the white ooze with specks, which would be consistent with the latter being seeds. The incoming fragment on the right was a direct body blow to Pod 2.

That incoming fragment missed Pod 1, missed the root of Pod 2, and went directly through the length of the body of Pod 2.
There is also evidence of periodic secondary hits on broken open pods, that result in cavitation.
In a cavitation impact, the material from the cavity typically becomes part of an "ejecta blanket". The white ooze and ejecta from all 3 sources likely accounts for the much lighter appearance of the carbonaceous chondrite relative to ordinary chondrites.
The white color of the pod contents indicates it may be calcium based. The pods appear to concentrate calcium in a form that makes it available to ambient life forms, while also spreading the seed of the pod across the land. The molecular biology implications of that are huge.
Calcium is arguably the most important element in multicellular life (Ch. 5).
At a sub cellular level, movement of the calcium ion Ca++ into and out of the cytoplasm functions as a signal for many cellular processes. The average adult human body contains 1.3 kg of calcium of which 99% is stored in the bone pantry, and routinely moved between the extracellular fluid, under endocrine control. Virtually every endocrine mediates its actions, in whole or in part, by moving calcium between bone and extracellular fluid (list in Ch. 5).
Intracellular cytosolic concentrations of Ca++ are kept low relative to extracellular concentrations. This concentration gradient drives Ca++ into the cell when Ca++ channels transiently open, which in turn activates Ca++ responsive proteins. Ca++ is required for function of nerves, muscles, secretions by cells, and cell growth and division pathways (population density management pathways).
Calcium is also the bridge to non physiological life. The calcium ion wave in the brain is what underlies the traveling direct electric current required for consciousness and memory formation. Using an opposing electric current, or magnetic clamping of this electron flow, results in loss of consciousness and memory formation (Dr. Robert Becker, "The Body Electric", 1985, p. 238). Dr. Becker demonstrated that this could be used to anesthetize animals.
UNDER CONSTRUCTION
IMPORTANT COPYRIGHT NOTICE: Copyright © 2012 - 2026 Mark J. Zamoyski. All rights reserved. No part of this website may be reproduced, scanned, stored, introduced into a retrieval system, or distributed in any printed or electronic form.

{kind=link}